

Гулин М.Б., Иванова Е.А.1
Цель работы – анализ средообразующей роли колебаний уровня Чёрного моря, происходящих в Голоцене под воздействием климатических факторов, на формирование современного черноморского шельфа: локальных мезомасштабных местообитаний в прибрежной зоне и ассоциированных с ними поселений зообентоса. Также, представляется важным сравнительное рассмотрение постгляциальной истории прибрежных экосистем Чёрного и Белого морей ввиду разнонаправленных трендов изменения их береговых линий.
Исследования последних лет позволили установить, что, по крайней мере, со средины 19-го века наблюдается устойчивое повышение уровня Мирового океана [http://www.climate.org/topics/sea-level/index.html#sealevelrise]. Скорость такого подъёма имеет практически линейный характер. За последние 100 лет поверхность Океана поднялась примерно на 20 см. Количественные оценки для более ранних эпох не так однозначны.
В Чёрном море скорость перманентного подъёма уровня вод в период 1876–1991 гг. составила 0,13 – 0,37 см/год [Коников, 2010]. Напротив, современные тектонические движения в черноморском регионе имеют противоположную по последствиям тенденцию: береговая линия крымского побережья за последние 10 миллионов лет поднялась на 10 метров [Фокина, 2006]. Однако, эта величина на три порядка меньше обратных процессов, происходящих в Голоцене – изменений водного баланса моря и осадконакопления, поэтому может не учитываться в оценках современной (кратковременной) геоэкологической эволюции черноморского бассейна.
Значимость описанных явлений представляется достаточно высокой и имеет в ряде случаев практическое значение. Так, например, в последние годы в Крыму участились оползни и обвалы в береговой зоне, повреждения прибрежной инфраструктуры.
К настоящему времени в Чёрном море экологическое значение поглощённых морем прибрежных зон практически не изучено. Вместе с тем, например, реликтовые участки устьевых и предустьевых речных систем на современном морском шельфе способны, вероятно, формировать особые условия обитания морских донных организмов. Речные бассейны, как правило, являются аккумуляторами осадочного материала, содержащего органическое вещество (ОВ) и после затопления их морем продолжают его накапливать за счёт естественного понижения дна в пределах своего батиметрического контура.
Впервые описание и целенаправленные исследования палеорусел рек Чёрная и Бельбек, а также их общей реликтовой дельты были выполнены сотрудниками ИМБИ РАН (в прошлом ИнБЮМ) в 2006–2007 гг. [Гулин, Коваленко 2010]. Прежде всего, были проведены детальные исследования батиметрических характеристик указанных мезоморф шельфа. Было установлено, в частности, что палеорусло реки Чёрная пролегает в северо-западном-западном направлении от кутовой части Севастопольской Бухты к кромке шельфа, оно довольно извилисто и имеет длину не менее 45 км. Палео-Бельбек, наоборот, отличается меньшей извилистостью и скорее похож на подводный канал. Реликтовое русло реки Бельбек простирается в направлении с востока на запад, практически строго вдоль широты 44°40´ с.ш. Обнаружено также, что реки Чёрная и Бельбек ранее сливались в единое русло или устье (рис. 1). Местоположение этого слияния находится на горизонте 86–89 м современного черноморского шельфа.
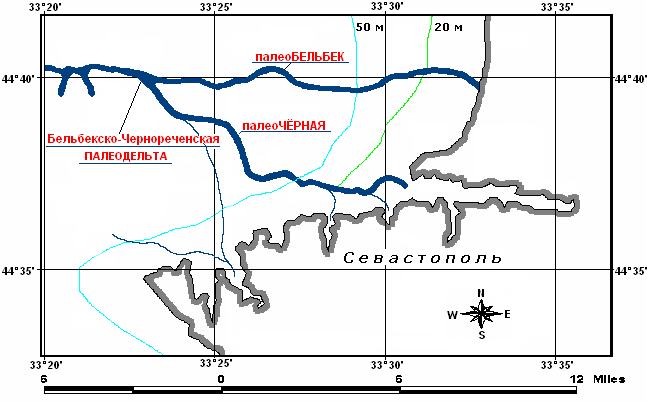
Рис. 1. Палеорусла и палеодельта рек Чёрная и Бельбек
Исследования конфигурации затопленного русла р. Чёрная на поперечных гидроакустических разрезах показали, что реликтовое ложе реки, причём как в пределах Севастопольской Бухты, так и на прилегающем взморье, имеет в сечении коробчатую форму с крутыми береговыми склонами и практически совершенно плоским коренным руслом.
Особенности профилей дна позволяют предположить, что затопленное морем корневое русло Чёрной ранее имело сечение скорее каньонного типа, но теперь покрыто мощным (более 40 м) слоем иловых отложений.
Поверхностные донные осадки в ложбине палеореки Чёрная значительно обогащены органическим веществом (ОВ), причём концентрация органики, в том числе её лабильных форм, в палеорусле многократно превышает таковую на прилегающих к нему участках дна (рис. 2). Последнее подтверждает, что палеорусла, как своеобразные углубления на дне шельфа, могут становиться “ловушкой” для оседающих из водной толщи взвесей.
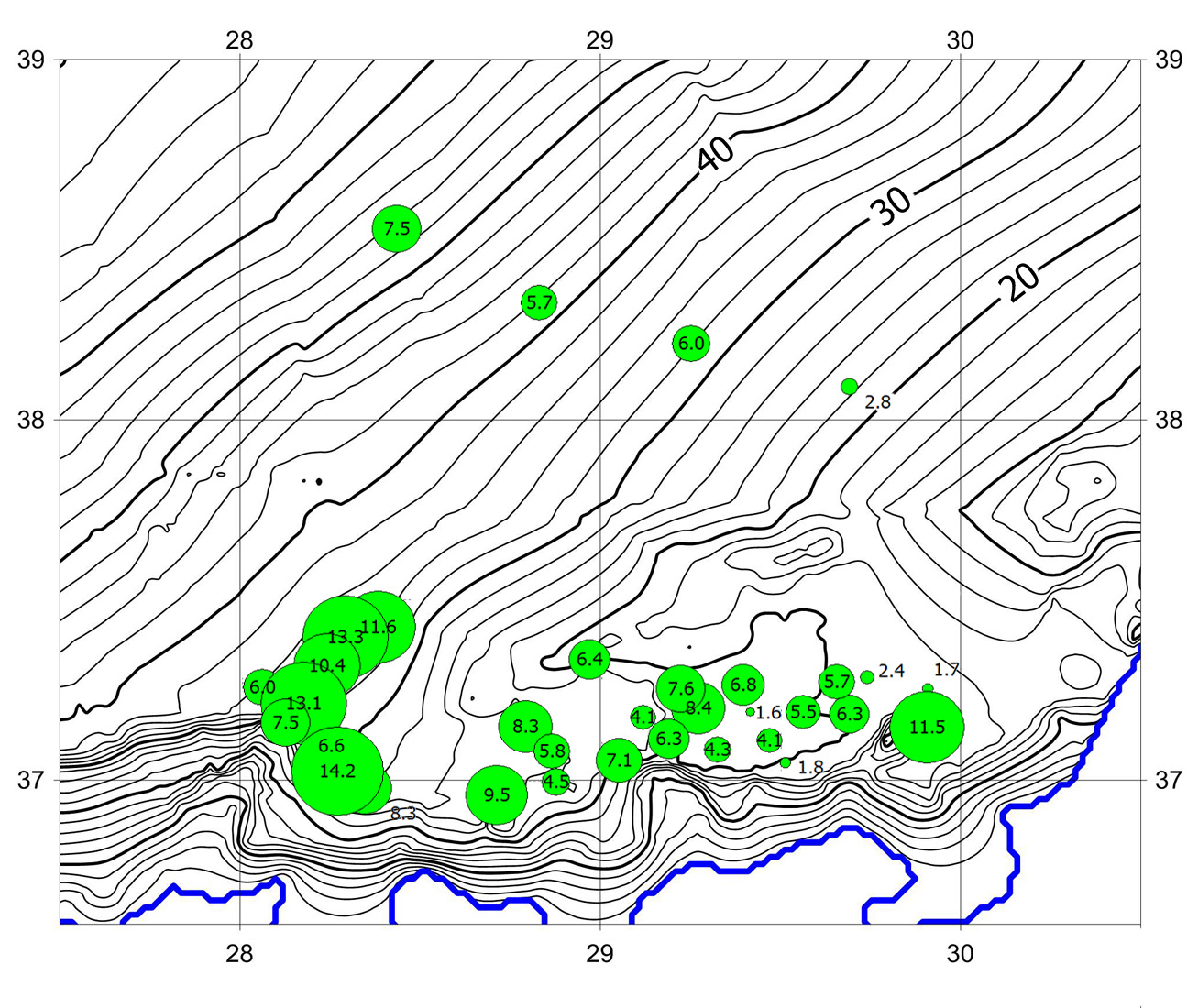
Рис. 2. Распределение легкоразлагающегося органического вещества
в палеорусле р. Чёрная (сжигание при +180 °С, мг/г сухого вещества)
Характер осадков палеорусла р. Чёрная, слагающих ложбину современного морского дна, показывает, что здесь накапливаются восстановленные, сульфидные илы и только на склонах самый верхний слой донных отложений может быть окисленным. Так, для поверхности донных осадков, залегающих непосредственно в ложе палео-Чёрной, характерны отрицательные показатели окислительно-восстановительного потенциала (Eh, мВ), тогда как на обоих склонах затопленной части реки – северном и южном – Eh имеет положительные значения. Соответственно, данный экотоп должен иметь выраженный гипоксический тип кислородного режима, определяющий специфику состава, обилия и распределения обитающих здесь гидробионтов-аэробов [Заика, Коновалов, Сергеева, 2011; Middelburg, Levin, 2009] и их функциональной активности.
Установлено, что ровное илистое дно палео-Чёрной повсеместно изрыто норами достаточно крупных представителей бентосной инфауны. При этом отверстия таких ходов были найдены лишь на вершинах гребней волновой ряби. Напротив, во всех случаях они отсутствовали в ложбинах между гребнями. Можно предположить, что указанная особенность распределения макрофауны является адаптацией, одним из способов избегания бентосными животными гипоксии и заморов.
Из всего спектра выявленных количественных показателей развития зообентоса в гипоксической бентали палеорусла реки Чёрная, наибольший интерес представляет сравнительный анализ параметров донной фауны в различных частях градиентного пространства реликтового фрагмента исследуемой речной системы – непосредственно в его ложе, а также на сопряжённых участках шельфа, ранее являвшихся береговыми склонами реки. Оказалось, что наиболее богатый видовой состав зообентоса характерен для береговых участков палеореки, а не его русла (рис. 3). На глубинах 16 и 74 м плотность поселений бентоса на реликтовых склонах превышала численность обитателей ложа палеорусла в 9 и 3 раза, соответственно. На мелководье (гл. 16 м) высокие количественные показатели развития бентофауны на береговых склонах были обусловлены, главным образом, высокой численностью Crustacea. На больших глубинах доминировали полихеты и немертины.
Аналогичные результаты, вернее, даже значительно больший гипоксический эффект наблюдался во внутренней акватории лимана Донузлав (северо-западный Крым). Было найдено, что в ложе данного полузамкнутого водоёма, практически на всём его протяжении, кислород в придонной воде отсутствует. Существенные величины [О٢] были зарегистрированы лишь на береговых склонах: 7,26–7,74 мг/л (глубины дна 3,6–6,7 м) у западного побережья и 7,59 мг/л (дно – 4,5 м) у противоположного восточного берега.
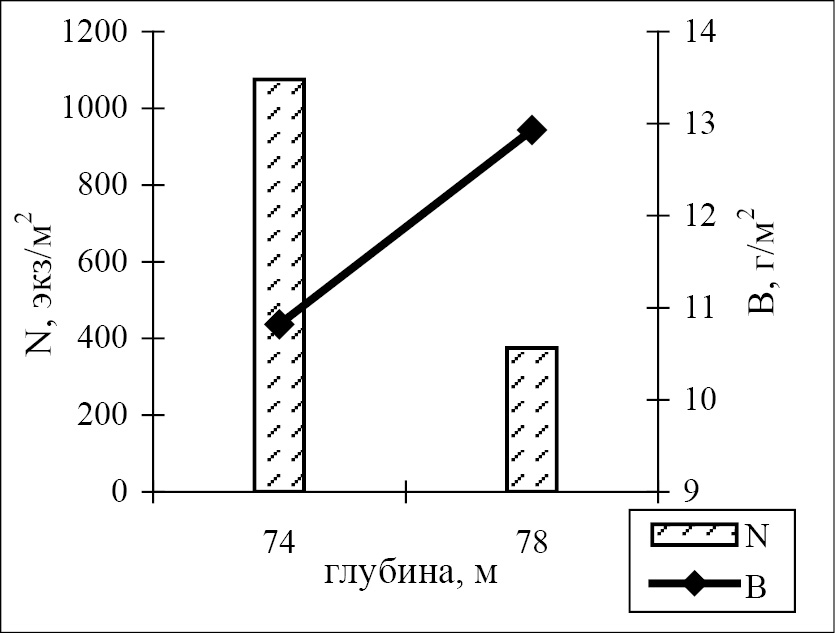
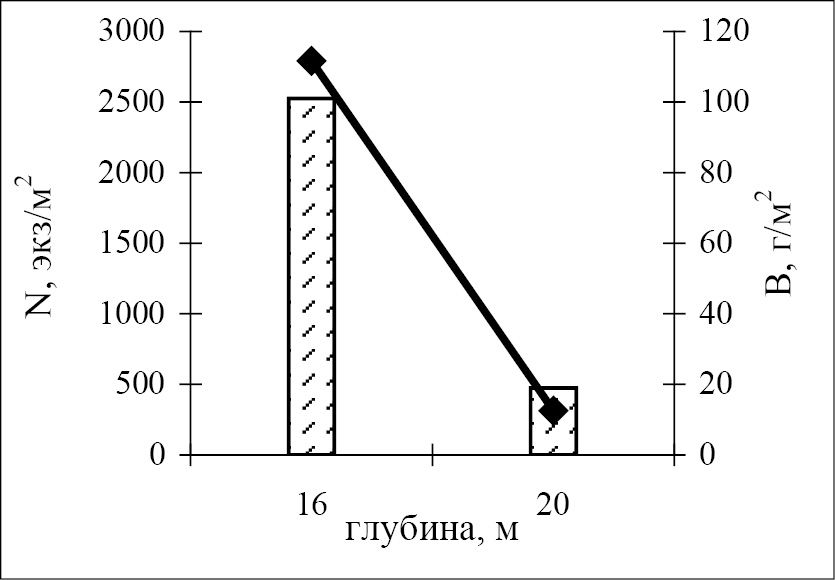
Рис. 3. Количественные параметры зообентоса в тальвеге (20 и 78 м)
и на береговых склонах (16 и 74 м, соответственно) на двух трансектах
через затопленное русло р. Чёрная; N – численность, B – биомасса
Анализируя в целом биологические параметры, характерные для ложа палеорусла р. Чёрная, следует отметить, что общее экологическое состояние бентосного сообщества в данном биотопе является угнетённым по сравнению с прилегающими осадками шельфа. Можно предположить, что данное явление связано с лимитирующими абиотическими факторами: повышенным содержанием ОВ, падением, как следствие его микробной минерализации, величины окислительно-восстановительного потенциала в донных осадках, дефицитом кислорода и сероводородным заражением на границе «придонный слой вод – поверхность дна». Можно констатировать, что обнаруженные гипоксические свойства исследуемого локального биотопа черноморской бентали прежде всего ответственны за обнаруженную обеднённость фауны макрозообентоса. Это согласуется с работами других авторов, показавших на иных локальных или региональных морских объектах, что гипоксия снижает плотность и биомассу бентоса [Levin, 2003; Quiroga E., 2005].
Отметим также, что этот достаточно очевидный факт несколько контрастирует с другим явлением – повышенным запасом органического вещества в ложбине палеорусла, которое, напротив, могло бы стать благоприятным трофическим ресурсом, привлекающим донных и демерсальных гидробионтов. Тем не менее, в сумме оба эти фактора сосуществуют и это позволяет причислить затопленные палеорусла на шельфе, как градиентные гипоксические биотопы, к единой общности прибрежных, сформировавшихся в постледниковый период локальных кислород-дефицитных местообитаний морских водоёмов – меромиктических лиманов Белого моря, районов газовых сипов и т.п., где также постоянно или периодически сосуществуют высокая трофность осадков с острым недостатком кислорода и токсичным сероводородным заражением.
Литература
- Гулин М.Б., Коваленко М.В. Палеорусла рек Чёрная и Бельбек на шельфе юго-западного Крыма – новый объект экологических исследований // Морской экологический журнал, 2010. – 9, №1. – С. 23–31.
- Коников Е.Г., Лиходедова О.Г. Глобальные и региональные факторы колебаний уровня Чёрного моря как основа геодинамической модели береговой зоны // Геология и полезные ископаемые Мирового океана, 2010. №1. – С. 84–93.
- Фокина Н.А. Природные факторы процессов абразии // Культура народов Причерноморья, 2006. №79. – С. 29–33.
- Заика В. Е., Коновалов С.К., Сергеева Н.Г. Локальные и сезонные явления гипоксии на дне севастопольских бухт и их влияние на макробентос // Морской экологический журнал. №3, Т. X, 2011. – С. 15 – 25.
- Levin L.A. Oxygen minimum zone benthos: adaptation and community response to hypoxia // Ann. Rev. Oceanogr. Mar. Biol, 2003. №. 41. – P. 1–45.
- Quiroga E., Quinones R., Palma M., Sellanes J., Gallardo V. A., Gerdes D., Rowe G. Biomass size-spectra of macrobenthic communities in the oxygen minimum zone off Chile // Estuar. Coast. Shelf S, 2005. №62. – P. 217–231.
- Middelburg J. J., Levin L. A. Coastal hypoxia and sediment biogeochemistry // Biogeosciences, 2009. №6. – P. 1273–1293.
1 Гулин М.Б., Иванова Е.А. – ФГБУН ИМБИ, Севастополь.