

Шатилович А.В.1, Мыльников А.П.2, Быкова С.В.3
Вечная мерзлота Арктики является уникальным криобанком генетических ресурсов. Многолетнемёрзлые толщи содержат значительное таксономическое разнообразие организмов, сохранивших жизнеспособность после сотен тысяч и миллионов лет криптобиоза в условиях отрицательных температур, отсутствия кислорода и доступной воды [Gilichinsky&Rivkina, 2011]. В мерзлоте, в числе прочих, обнаружены жизнеспособные гетеротрофные протисты: голые амёбы, жгутиконосцы и инфузории. Способность к образованию криптобиотических стадий, или цист покоя, позволяет почвенным протистам выживать при самых неблагоприятных условиях и расселяться на значительные территории. Описаны случаи сохранения цистами жизнеспособности после длительного (десятки лет) криптобиоза в сухих почвенных образцах. В 30-х годах прошлого века об обнаружении жизнеспособных цист простейших в мерзлых толщах Забайкалья сообщал Каптерев [1936, 1938]. Из образцов гренландского льда были выделены цисты инфузории Colpoda steinii и амебы Vahlkampfia sp., сохранившие способность к эксцистированию после нескольких столетий криоконсервации [Marquardt et al., 1966]. Проведенные нами исследования показали, что почвенные простейшие способны сохранять жизнеспособность в многолетнемёрзлых отложениях севера Якутии в течение десятков и сотен тысяч лет.
Материал для палеобиологических исследований был получен в ходе экспедиций «Берингия», проводившихся с 1990 по 2015 г. на приморских низменностях восточного сектора Арктики от дельты Лены до низовьев Колымы в области сплошного распространения вечной мерзлоты. Образцы отложений отбирали из керна основных позднекайнозойских горизонтов в тундровой зоне на побережье моря Лаптевых (мыс Быковский) и Восточно-Сибирского моря (устья рек Хрома, Бол. Куропаточья, Бол. Чукочья), а также в удаленных от моря районах (верховья р. Бол. Хомус-Юрях и среднее течение р. Бол. Чукочья). Диапазоны среднегодовой температуры пород от –9°С (в депрессиях) до –12°С (на водоразделах), максимальной глубины сезонного оттаивания суглинистых почв водоразделов – от 40 до 60 см. Образцы погребенных почв и ископаемых нор отбирали в современной зоне северной тайги в восточном борту Колымской низменности из обнажений позднеплейстоценового ледового комплекса Омолоно-Анюйской едомы (Станчиковский яр и Дуванный яр). Среднегодовая температура пород составляет здесь –5…–6°С, а глубина сезонного оттаивания достигает 80 см. Погребенные почвы в отложениях ледового комплекса представлены торфяником и профилями перегнойноторфянистой глеевой почвы [Губин, 1994]. Отбор образцов и их транспортировку осуществляли с соблюдением необходимого температурного режима и требований стерильности согласно методикам, многократно апробированным лабораторией криологии почв ИФХиБПП РАН при микробиологических исследованиях мёрзлых толщ [Shi et al., 1997].
Проанализировано более 300 образцов многолетнемёрзлых отложений разного возраста и генезиса, погребённых почв и нор ископаемых грызунов. Жизнеспособные цисты простейших обнаружены в эпикриогенных осадках голоценовых аласов и в синкриогенных отложениях позднеплейстоценового ледового комплекса. В керне скважин простейшие чаще (в 60% случаев) встречались в голоценовых осадках в первых метрах от дневной поверхности. Наибольшая глубина, с которой выделены жизнеспособные простейшие, равна 30 м. Простейшие обнаружены в 20% исследованных образцов ледового комплекса. В погребенных почвах и материале ископаемых нор встречаемость была значительно выше: 80% и 100%, соответственно. Длительность криптобиоза простейших в кровле вечной мерзлоты не превышает нескольких тысяч лет, а самые древние, единичные находки относятся к среднему плейстоцену – 200–300 тыс. лет. Комплексы жизнеспособной микрофауны, выделенной из мерзлоты, составляют 13 видов инфузорий (Рис.1) и 26 видов жгутиконосцев (Рис. 2). Основу видового разнообразия ископаемых простейших составляют эврибионтные виды, широко представленные в современных разнотипных водных и почвенных экосистемах.
Ряд факторов позволяет нам исключить возможность проникновения цист простейших в многометровые мёрзлые толщи из расположенной выше современной тундровой почвы. Глубина сезонного оттаивания в районах исследования может достигать 80 см, ниже этого уровня, в многолетнемёрзлых, прочно сцементированных льдом отложениях позднеплейстоценового ледового комплекса, влияние внешних факторов резко ограничено, отсутствуют водоносные горизонты и инфильтрация. Термодиффузия и миграция цист с плёнками незамёрзшей воды, толщина которых не превышает нескольких нанометров, невозможна. Наличие мощных жильных льдов служит доказательством того, что вмещающие их породы являются синкриогенными и никогда не оттаивали. Возраст биоты в синкриогенных толщах соответствует возрасту осадочных пород [Гиличинский и др., 1989].
Подавляющее большинство обнаруженных жизнеспособных протистов получено из материала, обогащенного растительными остатками: погребенных эпигенных почв, древних нор грызунов, растительного детрита.
Криосинлитогенное почвообразование в позднем плейстоцене характеризовалось регулярным поступлением на поверхность небольших доз органо-минерального осадка и последующим воздействием на него комплекса почвенных процессов в резко континентальных климатических условиях развития тундростепных биоценозов [Губин, 2002]. Присутствие профилей погребенных почв в толще криопедолита указывает на наличие на стадии МИС-3 периодов снижения активности поступления минерального осадка на поверхность и усиления гидроморфизма. Стабилизация дневной поверхности в течение, как минимум, сотни лет, формирование органогенных горизонтов и стабильное увлажнение в летний период создавало предпосылки для образования в почвах развитых и устойчивых биоценозов, включающих и более богатую фауну протистов.
К особым палеэкологическим объектам относятся ископаемые норы сусликов, принадлежащих одному из видов подрода Urocitellus [Губин и др., 2003]. Норы приурочены к толщам криопедолита каргинского возраста (стадия МИС-3) и связаны с периодами суровых климатических условий при широком распространении равнинных тундростепных ландшафтов. Занесенные с поверхности вместе с растительным материалом и шерстью крупных животных мамонтовой фауны, цисты простейших сохранялись в сухих, хорошо аэрируемых и защищенных от резких перепадов температур камерах, и относительно быстро переходили в мерзлое состояние. Наши исследования показали, что сообщества жизнеспособных простейших, выделенных из многолетнемерзлых позднеплейстоценовых погребенных почв и нор, отличаются более высоким видовым разнообразием и численностью по сравнению с вмещающими отложениями ледового комплекса. Предполагается, что растительные остатки являлись не только естественным убежищем для цист и спор древних организмов, но и выполняли криопротекторные функции.
Работа выполнена в рамках Госзаданий АААА-А18-118013190181-6 и при поддержке гранта РФФИ №17-04-01397.
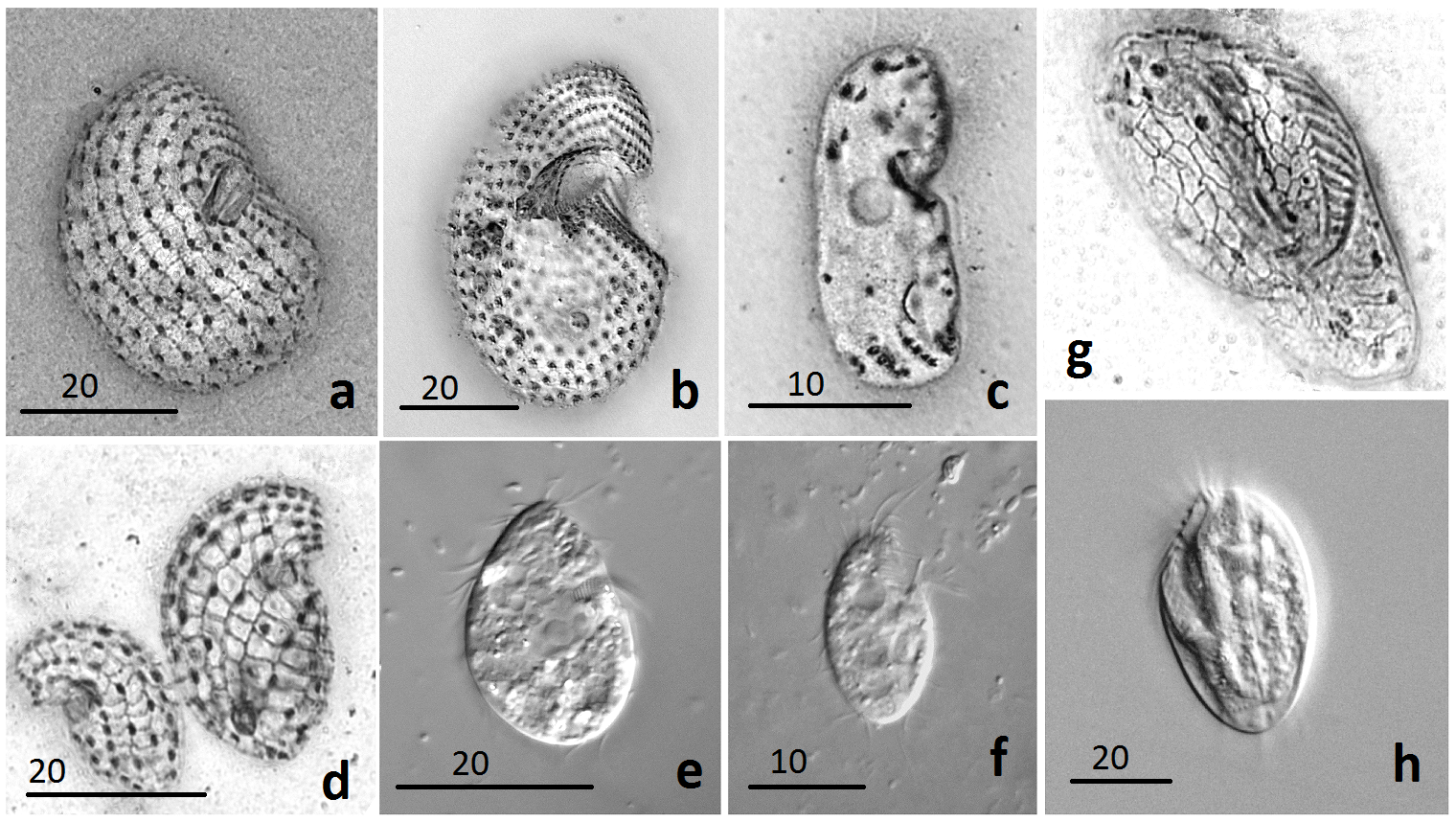
Рис. 1. Жизнеспособные инфузории, выделенные из многолетнемёрзлых
отложений северной Якутии: a – Colpoda inflata; b – Colpoda cucculus;
c – Drepanomonas sphagni; d,e – Colpoda steinii; f – Cyrtolophosis mucicola;
g,h – Euplotes moebius. Scale bars: 10 µm (c,f), 20 µm (a,b,d,e,g,h)
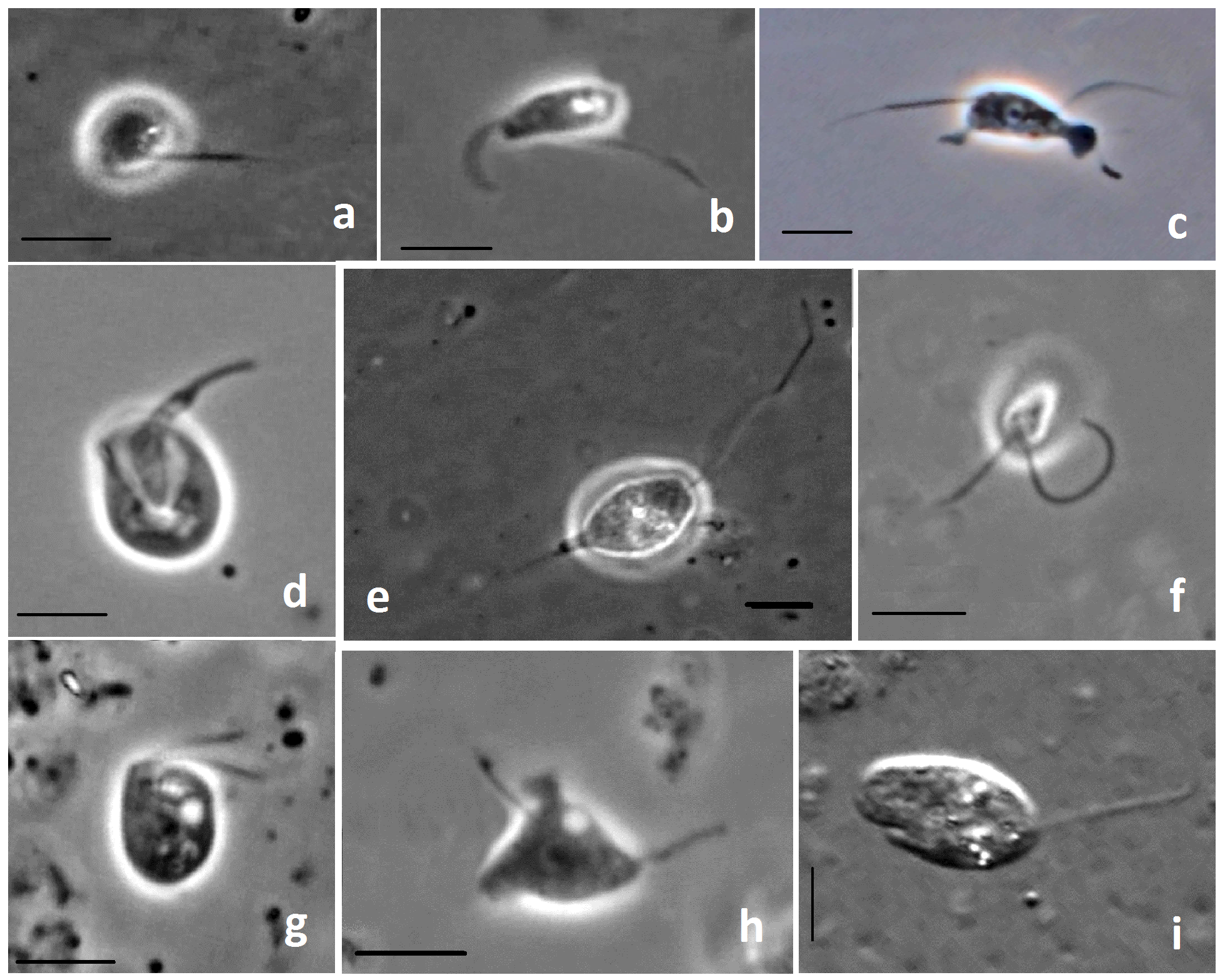
Рис. 2. Жизнеспособные гетеротрофные жгутиконосцы, выделенные из многолетнемёрзлых отложений северной Якутии: a – Allapsa sp.; b – Neobodo designis; c – Paracercomonas krugeri; d – Apusomonas aff. australiensis; e – Cercomonas directa;
f – Neoheteromita globosa; g – Goniomonas truncata; h – Paracercomonas proboscata;
i – Eocercomonas exploratorii. Scale bars – 5 µm
Литература
- Gilichinsky D., Rivkina E. Encyclopedia of Earth Sciences Series. B.: Springer, 2011. – P. 726–732.
- Marquardt W.C., Wang G.-T., Fennell D.I. Preservation of Colpoda stenii and Vahlkampfia sp. from ice tunnel in Greenland // Trans. Amer. Microscop. Soc., 1966, vol. 85, No. 1, – Р. 152–156.
- Shi T., Reevs R., Gilichinsky D., Friedman E.I. // Microbiol. Ecol. 1997. №33. – P. 169–179.
- Гиличинский Д.А., Хлебникова Г.М., Звягинцев Д.Г., Федоров-Давыдов Д.Г., Кудрявцева Н.Н. // Изв. АНСССР. Сер. геол. 1989. №6. – С. 114–126.
- Губин С.В. Позднеплейстоценовое почвообразование на приморских низменностях севера Якутии // Почвоведение,1994, №8. – С. 5–14.
- Губин С.В. Педогенез – составная часть механизма формирования отложений позднеплейстоценового ледового комплекса // Криосфера Земли, 2002, т. VI, №3. – С. 82–91.
- Губин С.В., Занина О.Г., Максимович С.В. и др. Реконструкция условий формирования отложений ледового комплекса по результатам изучения позднеплейстоценовых нор грызунов // Криосфера Земли, 2003, т. VII, №3. – С. 13–22.
- Каптерев П.Н. Об анабиозе в условиях вечной мерзлоты // Изв. АН СССР. Сер. биол., 1936, №6. – С. 1073–1088.