

Саввичев А.С.1, Летаров А.В., Лунина О.Н.,
Веслополова Е.Ф., Демиденко Н.А.2,
Кокрятская Н.М.3, Краснова Е.Д.4, Горленко В.М.
Меромиктические водоемы, имеющие вертикальную стратификацию по температуре и содержанию солей, являются классическими объектами исследований лимнологов, биогеохимиков, микробиологов [Кузнецов, 1970]. В водной толще меромиктических водоемов в слое хемоклина развивается сложное многослойное микробное сообщество, участвующее в преобразовании соединений углерода, серы, железа, марганца [Ivanov et al., 2001; Pimenov et al., 2003; Саввичев с соавт., 2004, Crowe et al., 2010; Galand et al., 2012; Wand et al., 2006].
Озеро Трехцветное (66° 35.53’ N, 32° 59.97’ E) – меромиктический водоем классического типа. Это небольшое (150 м в ширину и 340 м в длину) озеро с максимальной глубиной 7,5 м расположено в 12 км от ББС МГУ. Название «Трехцветное» этому озеру дано за отчетливое различие в окраске трех слоев водной толщи [Пантюлин, Краснова, 2011]. Актуальность исследования водоемов, отделенных от основного морского бассейна, связана, прежде всего, с необходимостью прогноза негативных последствий сероводородного заражения в искусственно замкнутых морских акваториях, появившихся в результате строительства дамб, плотин, приливных электростанций и т.д.
Общая численность и биомасса микроорганизмов. Микроскопирование окрашенных препаратов показало крайне неравномерное распределение микроорганизмов в водной толще оз. Трехцветное (рис. 1).
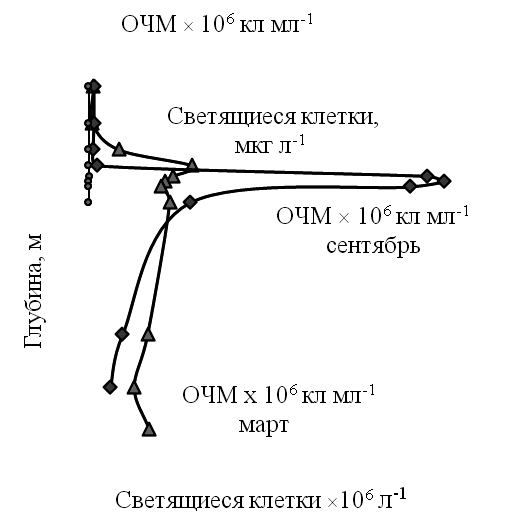
Рис. 1. Общая численность микроорганизмов (ОЧМ) водной толщи оз. Трехцветное
Величина общей численности микроорганизмов (ОЧМ) была относительно высокой в верхнем кислородсодержащем водном слое (ср. 3,0 млн кл мл-1). В высокоплотном «густо-зеленом» слое хемоклина величина ОЧМ достигала крайне высоких значений − до 60 млн кл мл-1 (слой 1,8 м) в зимний сезон 2013 г. и до 210 млн кл мл-1 (слой 2,0 м) в сентябре 2014 г. Средний объем клеток бактерий в аэробной водной толще составлял 2,5 мкм3, а в «густо-зеленом» слое достигал 2,0 мкм3. Соответственно величина микробной биомассы в аэробном слое составляла 0,5−0,7 мг л-1, а в слое максимальной плотности 42 мг л-1.
Продукция фотосинтеза и интенсивность темновой СО٢ ассимиляции в водной толще озера. Для меромиктических водоемов и для оз. Трехцветное, в частности, характерно два типа фотосинтеза: оксигеный, осуществляемый эукариотическими водорослями и цианобактериями в поверхностном водном слое и аноксигенный, осуществляемый аноксигенными фототрофными бактериями (АФБ) в глубинном слое. В августе в аэробной зоне активность фотосинтеза была зарегистрирована в поверхностном слое до 1,8 м включительно (рис. 2).
Интенсивность оксигенного фотосинтеза составляла 12−24 ммоль С л-1 сут-1. На нижней границе аэробной зоны интенсивность фотосинтеза резко снижалась. Второй, более выраженный, чем первый, пик световой фиксации углекислоты проявлялся в верхней части сероводородсодержащего, «густо-зеленом» слоя, на горизонтах 1,9 и 2,0 м (до 240 мкмоль С л-1 сут-1). При дальнейшем погружении интенсивность фотосинтеза резко падает и, в минимальных значениях обнаруживается даже в придонном горизонте (до 0,8 мкмоль С л-1 сут-1).
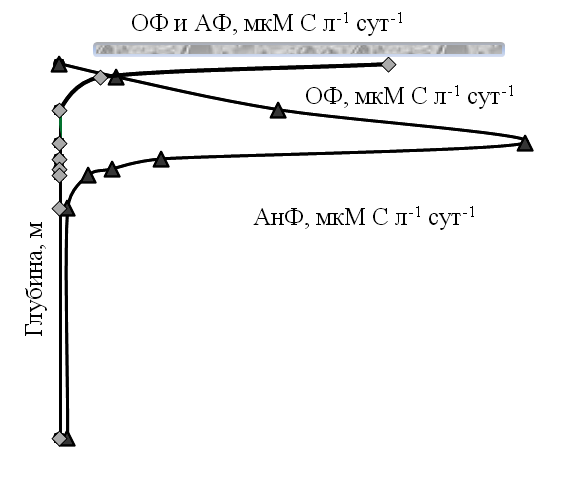
Рис. 2. Интенсивность оксигенного (ОФ) фотосинтеза и аноксигенного (АнФ)
фотосинтеза в водной толще оз. Трехцветное в зимний и летний сезоны
Темновая ассимиляция углекислоты (ТАУ) является суммарным показателем, включающим как интенсивность гетеротрофного карбоксилирования, так и автотрофное потребления углекислоты при хемосинтезе. В аэробном водном слое величина ТАУ является характеристикой активности фототрофного и гетеротрофного бактериопланктона (3−6,5 мкмоль С л-1 сут-1 в августе, (рис. 2). В этом водном слое хемоавтотрофные процессы протекают слабо, из-за недостатка неорганических восстановителей. В зеленом слое величина ТАУ скачкообразно возрастает до 27,3 и 47,8 мкмоль С л-1 сут-1 в сентябре. На нижележащих водных горизонтах величина ТАУ плавно падает от редокс-зоны к придонному слою (до 8,0−14,0 мкмоль С л-1 сут-1 в сентябре). Процесс ассимиляции углекислоты осуществляется как за счет анаэробных гетеротрофных микроорганизмов, так и за счет анаэробных хемоавтотрофов: гидрогенотрофных сульфатредукторов и метаногенов.
Аноксигенные фототрофные бактерии оз. Трехцветное. Из накопительного флакона, с глубины 1 м, была выделена культура ЗСБ штамм GrTcv12. Клетки были мелкие, неподвижные, округлые или овальные, размером 0,1–0,15 х 0,2–0,3 мкм, содержали газовые вакуоли и могли образовывать короткие и длинные (более 20 клеток) изогнутые нити. Оптимальная соленость для роста штамма GrTcv12 составляла 5–10 г/л NaCl. Фотосинтетическими пигментами были Бхл d и каротиноид хлоробактин. Из колоний, выросших на агаризованной среде при посеве пробы воды с глубины 1,5 м, были выделены пурпурные серные бактерии (ПСБ) AmTсv13. Это были неподвижные округлые клетки 1–1,7 мкм в диаметре, содержащие газовые вакуоли и капли серы. Культура хорошо вырастала при солености 10–35 г/л NaCl, оптимальная соленость составляла 10 г/л. По данным сиквенса 16S рДНК штамм AmTсv13 оказался наиболее близок к штамму Thiocapsa sp. AmPS10 (номер Генбанка КС702856), выделенному из озера Кисло-Сладкое осенью 2010 г. [Лунина с соавт., 2014], и имеет с ним 99% сходства.
Процесс бактериальной сульфатредукции. Проведенные радиоизотопные эксперименты показали, что процесс сульфатредукции (СР) начинает реально проявляться на горизонте 1,75 м, соответствующему верхнему горизонту редокс зоны. Ниже интенсивность СР возрастает и достигает максимально высоких значений (19−34 мкмоль S л-1 сут-1) в «густо-зеленом» водном горизонте 2,0−2,1 м. Именно на этом горизонте процесс образования свежего ОВ за счет аноксигенного фотосинтеза протекает наиболее активно. Ниже 2,1 м интенсивность СР снижается: до 15 мкмоль S л-1 сут-1 на горизонте 2,5 м и до 10 мкмоль S л-1 сут-1 на горизонте 6,0 м. Изотопный состав углерода ОВ взвеси в поверхностном слое водоема оказался близок к значениям, характерных для пресноводной взвеси, что указывает на влияние пресного стока из окружающих озеро заболоченных берегов.
Горизонт 1,5 м, который характеризуется локальным максимумов активности оксигенного фотосинтеза был обогащен тяжелым изотопом углерода ОВ (δ13CCорг. = –25,1‰) в составе новообразованного автохтонного фитопланктона. Увеличение доли тяжелого изотопа связано с фракционированием, осуществляемым оксигенными фототрофами, потребляющими преимущественно легкий изотоп карбонатного углерода [Пименов с соавт., 2008]. Начиная с горизонта 1,75 м и до 2,1 м наблюдается заметное облегчение изотопного состава углерода ОВ (δ13CCорг. = –28,8‰ – –32,9‰). В глубинном анаэробном водном слое величина δ13CCорг. варьирует от –25,4 до –26,0‰, а величина δ13C∑CО٢ от –8,9 до –9,2‰, при этом эффективность фракционирования составляет ∆ = 16,4−16,9‰. Облегчение изотопного состава минерального углерода в придонных горизонтах связано с интенсивными процессами анаэробной деструкции ОВ, образованного сообществом аноксигеных фототрофных, хемотрофных и метанотрофных микроорганизмов в вышерасположенном водном слое.
Среди многих прибрежных водоемов Кандалакшского побережья, имеющих ограниченную связь с основным морским бассейном, оз. Трехцветное выделяется наибольшей стабильностью гидрологического режима. В настоящее время абсолютная отметка уреза воды (замер 2014 года) составляет 0,85 м [Краснова с соавт., 2014]. Озеро отделено от Пеккелинской губы заросшей протокой, при этом поступление свежей морской воды является крайне редким событием, случающимся при совпадении сильного нагонного ветра с сизигийным приливом. Вероятно, что именно стабильность в стратификации приводит к существованию плотного «зеленого слоя». Плотные микробные слои, выделяющиеся и различимые за счет своего естественного красного или зеленого цвета, иногда получали название «бактериальных пластин» [Vila et al., 1998; Garcia-Gil et al., 1999; Camacho et al., 2001, Рогозин с соавт., 2010]. Было показано, что численность пурпурных серных бактерий в плотном слое оз. Махони (Mahoney), Канада превышает 108 кл мл-1 [Overmann, 1997]. Исследованное нами озеро Трехцветное отличалось тем, что фототрофное микробное сообщество плотного слоя было представлено только зелеными серными бактериями. Общая численность микроорганизмов в этом слое достигала 2,1 × 108 кл мл-1.
Авторы признательны коллегам из разных учреждений, принявших участие в летних и зимних исследованиях оз. Трехцветное Д.А. Воронову, Н.Л. Фроловой, С.В. Пацаевой, А.В. Харчевой, С.А. Забелиной, Г.Н. Лосюк. Работа выполнена с использованием финансовых средств грантов РФФИ 17-04-01263 и 16-05-00548.
Литература
- Краснова Е.Д., Демиденко Н.А., Пантюлин А.Н., Фролова Н.Л., Ефимова Л.Е., Широкова В.А. Термический и ледовый режимы реликтовых водоемов, отделяющихся от Белого моря // Динамика и термина рек, водохранилищ и прибрежной зоны морей : труды VIII Межд. научно-практической конф.: в 2 т. – М.: РУДН, 24–27 ноября 2014 г. – М. : РУДН, 2014. – С. 430−443.
- Кузнецов С. И. Микрофлора озер и ее геохимическая деятельность. – Л.: Наука, 1970. 440 с.
- Лунина О. Н., Саввичев А.С., Пименов Н.В., Кузнецов Б.Б., Горленко В.М. Аноксигенные фототрофные бактерии оз. Кисло-Сладкое (Кандалакшский залив Белого моря) // Микробиология. 2013. Т. 82. №6. –С. 815−832.
- Пантюлин А.Н., Краснова Е.Д. Отделяющиеся водоемы Белого моря: новый объект для междисциплинарных исследований. «Геология морей и океанов»: Материалы XIX Международной научной конференции (Школы) по морской геологии. Т. III. – М.: 2011. – С. 241–245.
- Пименов Н.В., Лунина О.Н., Прусакова Т.С., Русанов И.И., Иванов М.В. Биологическое фракционирование стабильных изотопов углерода на границе аэробных и анаэробных вод в меромиктических водоемах // Микробиология. 2008. Т. 77. №6. – С. 839–847.
- Рогозин Д.Ю., Трусова М.Ю., Хромечек Е.Б., Дегерменджи А.Г. Микробное сообщество хемоклина меромиктического озера Шунет (Хакасия, Россия) в период летней стратификации // Микробиология. 2010. Т. 79. №2. – С. 277–285.
- Саввичев А.С., Русанов И.И, Юсупов С.К., Пименов Н.В., Леин А.Ю., Иванов М.В. Биогеохимический цикл метана в прибрежной зоне и литорали Кандалакшского залива Белого моря // Микробиология. 2004. Т. 73. №4. – С. 540–552.
- Camacho A., Erez J., Chicote A., Florin M., Squires M.M., Lehmann C., Bahofen R. Microbial microstratification, inorganic carbon photoassimilation and dark carbon fixation at the chemocline of the meromictic Lake Cadagno (Switzerland) and its relevance to the food web // Aquat. Sci., 2001. – V. 63. – P. 91−106.
- Crowe S.A., O’Neill A.H., Katsev S., Hehanussa P., Haffner G.D., Sundby B., Mucci A., Fowle D.A. (2008) The biogeochemistry of tropical lakes: a case study from Lake Matano, Indonesia. Limnology and Oceanography 53, – P. 319–331.
- Galand P.E., Bourrain M., De Maistre E., Catala P., Desdevises Y., Elifantz H., Kirchman D. L., Lebaron P. Phylogenetic and functional diversity of Bacteria and Archaea in a unique stratified lagoon, the Clipperton atoll (N Pacific) // FEMS Microbiol. Ecol. 2012. №79. – Р. 203–217.
- Garcia-Gil L.J., Vicente E., Camacho A., Borrego C.M., Vila X., Cristina X.P., Rodriguez-Gonzalez J. Vertical distribution of photosynthetic sulphur bacteria linked to saline gradients in Lake El Tobar (Cuenca, Spain) // Aquat. Microb. Ecol., 1999, V. 20. – P. 299−303.
- Overmann J. Mahoney Lake: a case study of the ecological significance of phototrophic sulphur bacteria // Adv. Microb. Ecol. 1997. V. 15. – P. 251–288.
- Vila X., Abella C.A., Figueras J.B., Hurley J.P. Vertical models of phototrophic bacterial distribution in the metalimnetic microbial communities of several freshwater North-American kettle lakes // FEMS Microb. Ecology, 1998, V. 25. – P. 287−299.
1 Саввичев А.С., Летаров А.В., Лунина О.Н., Веслополова Е.Ф., Горленко В.М. – Институт микробиологии им. С.Н. Виноградского ФИЦ РАН, Москва.
2 Демиденко Н.А. – Государственный океанографический Институт им. Н.Н. Зубова, Москва.
3 Кокрятская Н.М. – Институт экологических проблем Севера УрО РАН, Архангельск.
4 Краснова Е.Д. – МГУ им. М.В. Ломоносова, биологический факультет, Москва.