

Регуляция секреции
Гормоны щитовидной железы – тироксин (T4) и трийодтиронин (T3) являются йодированными производными аминокислоты тирозина, имеющими в своей молекуле четыре (Т4) или три (Т3) атома йода. Они синтезируются в клетках фолликуляного эпителия (тиреоцитах) щитовидной железы в составе белка тироглобулина и в такой форме поступают во внутрифолликулярный коллоид. При секреторном стимуле (в основном под действием ТТГ) клетки фолликулов осуществляют эндоцитоз коллоида и его лизосомальныий протеолиз с высвобождением тиреоидных гормонов, которые за счет своей липофильности проникают через клеточную мембрану и поступают в системный кровоток.
Тироксин является малоактивным тиреоидным гормоном и слабо связывается с соответствующими рецепторами в тканях, поэтому часто рассматривается как прогормон трийодтиронина. У здорового человека секреция Т4 щитовидной железой в 5–10 раз превышает секрецию Т3. Трийодтиронин – существенно более активный гормон, способный дополнительно образовываться из тироксина в тканях-мишенях за счет отщепления атома йода дейодиназой определенного типа. Именно Т3 в основном определяет биологические эффекты тиреоидных гормонов.
Биосинтез и секреция гормонов щитовидной железы стимулируется тиреотропным гормоном (ТТГ) гипофиза. В свою очередь секреция ТТГ находится под положительным контролем тиролиберина (ТРГ38) гипоталамуса. Т3 и Т4 действуют на продукцию ТРГ и ТТГ по принципу отрицательной обратной связи (см. рис. 29). Повышение секреции тиреоидных гормонов ведет к снижению продукции ТРГ и ТТГ, снижение вызывает обратные процессы. Таким образом, секреция гормонов щитовидной железы поддерживается на относительно постоянном уровне. Решающим сигналом обратной связи является плазменная концентрация Т4. В клетках тиреотрофов аденогипофиза и нейронах гипоталамуса он превращается в более активный Т3. Трийодтиронин тормозит синтез ТТГ и снижает чувствительность тиреотрофов к тиролиберину39. Продукция тиролиберина и ТТГ регулируется также за счет наличия короткий обратных связей.
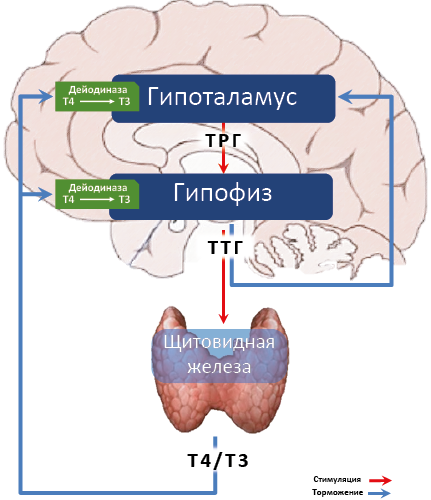
Рис. 29. Гипоталамо-гипофизарно-тиреоидная ось
Из-за короткого периода полувыведения и быстрого действия трийодтиронина (ЛС лейотиронин) пациентам с гипотиреозом (недостаточностью щитовидной железы) чаще назначают именно тироксин (ЛС Левотироксин натрия), который назначают в индивидуальной дозировке 1 раз в день. Назначение аналогов Т3 целесообразно только в случае неспособности организма самостоятельно преобразовывать Т4 в Т3.
Дефицит йода в организме может служить причиной развития диффузного нетоксического зоба – так называемого «эндемического зоба». Для него характерен пониженный уровень тиреоидных гормонов в крови. Эндемичны по дефициту йода:
• Алтай
• Кавказ
• Альпы
• Центральная Россия
• Средняя Азия и др.
Если дефицит йода затрагивал эмбриональный и детский периоды, может наблюдаться отставание в интеллектуальном развитии. Если дефицит йода не устранён, диффузный нетоксический зоб становится узловым нетоксическим, а затем – узловым токсическим.
Появление стимулирующих аутоантител к рецепторам ТТГ – причина развития диффузного тиреотоксического зоба (болезни Грейвса). Болезнь характеризуется бессонницей, экзофтальмом (смещение глазных яблок вперёд), раздражительностью и др. симптомами. Причина экзофтальма – пролиферация фибробластов, ретроорбитального (заглазного) пространства под действием тиреоидных гормонов.
При тиреотоксикозе назначают производные тиомочевины Пропилтиоурацил и Тиамазол, ингибирующие тиропероксидазу и, таким образом, нарушающие йодирование тирозиловых остатков тироглобулина. Пропилтиоурацил также угнетает превращение Т4 вТ3 на периферии. Перед операциями на щитовидной железе для угнетения синтеза и выделения Т3 иТ4 применяют йодиды калия или натрия в высоких дозах (160–180 мг).
Структура, биодинамика и механизмы действия
Тиреоидные гормоны являются йодированными производными тирозина, следовательно, для их нормального синтеза необходимо адекватное и регулярное поступление йода с пищей. Это отличает их от прочих сигнальных соединений.
ТТГ – гликопротеидный гормон семейства ФСГ40, состоит из двух цепей:
- α-цепи, общей для всех представителей семейства;
- β-цепи, специфичной для ТТГ.
Тиролиберин – трипептидный рилизинг-гормон гипоталамуса. Синтезируется путем выделения нескольких трипептидов из крупномолекулярного предшественника. Помимо стимуляции синтеза ТТГ, также повышает продукцию пролактина и действует в ЦНС как нейропептид.
Для Т3/Т4 известно несколько транспортных белков крови, в частности, тироксинсвязывающий глобулин (ТСГ) и транстиретин (тироксинсвязывающий преальбумин). ТСГ – основной белок, связывающий тиреоидные гормоны, в комплексе с ним транспортируется более 75% всех тиреоидных гормонов. Его сродство к тироксину в 4-5 раз больше, чем к трийодтиронину, поэтому концентрация свободных Т3 и Т4 в крови сопоставима, несмотря на непропорциональную продукцию щитовидной железой. Отщепление атома йода под действием дейодиназ является основным путем метаболизма гормонов щитовидной железы. В зависимости от положения дейодирования может происходить их тканевая:
- активация (по 5` положению с образованием из Т4 более активного Т3)
- инактивация (по 5 положению с образованием обратного Т3 и др. веществ)
В печени и других тканях катаболизма процесс на этом не останавливается и постепенно происходит полное удаление атомов йода.
Рецепторы
Рецепторы тиреоидных гормонов относятся к надсемейству ядерных рецепторов, адаптивных факторов транскрипции. В отсутствие гормона рецептор связан с регуляторной областью ДНК гормончувствительного гена и препятствует его транскрипции. После связывания Т3 или Т4 с рецептором снимается его репрессорное действие и происходит считывание информации.
Рецепторы ТТГ сопряжены с G-белками и располагаются на базальной части мембраны тиреоцитов. После связывания β-субъединицы ТТГ с рецептором происходит активация аденилатциклазной41 системы. Повышение уровня цАМФ увеличивает:
- транскрипцию гена тироглобулина;
- поступление йодидов в фолликулы щитовидной железы (транскрипцию транспортеров йодидов);
- йодирование тироглобулина (транскрипцию йодидпероксидазы);
- захват тиреоцитами коллоида со зрелым тироглобулином.
Рецепторы тиролиберина также относятся к надсемейству рецепторов, сопряженных с G-белками, но передают сигнал преимущественно по фосфатидилинозитольному42 пути с участием ионов Са2+.
Функции гормонов щитовидной железы
Функции гормонов щитовидной железы различны в раннем онтогенезе и у взрослых особей. К функциям тиреоидных гормонов относят:
Регуляцию роста и развития:
- программирование развития мозга;
- программирование архитектоники мозга;
- дифференцировка клеток мозга;
- синаптогенез;
- миелинизация;
Рост и развитие различных систем;
Обмен веществ:
- Стимуляция аэробного метаболизма;
- Регуляция метаболизма белков, липидов и углеводов;
- Термогенез;
- Регуляция транспорта ионов;
Стимуляция:
- Сердечной деятельности;
- Дыхания;
- Активности ЦНС;
Регуляция репродуктивной ф-ции;
Информационные ф-ции:
↑ СТГ;
↑ соматолиберина;
↑ Na+-уретических пептидов;
↑ гастрина;
↑ ГСПГ43.
Регуляция общего развития
В раннем онтогенезе тиреоидные гормоны играют определяющую роль в процессах нормального развития. Они влияют на дифференцировку и рост тканей, развитие ЦНС (стимулируя дифференцировку разных типов клеток мозга, синаптогенез и миелинизацию нейронов). Гормоны щитовидной железы стимулируют увеличение размеров организма, действуя на ряд компонентов соматотропной оси (см. раздел Соматотропная ось). У низших позвоночных, например, амфибий, тиреоидные гормоны стимулируют метаморфоз.
Влияние на мозг
Основной функцией Т3 и Т4 во время внутриутробного периода развития является дифференцировка тканей, прежде всего нервной. Для нормального развития мозга плода необходимо достаточное количество тироксина, который в нервной ткани частично превращается в трийодтиронин. Первоначально необходимый уровень Т4 обеспечивается материнским организмом, после середины беременности – щитовидными железами и матери, и плода. В эмбриогенезе тиреоидные гормоны способствуют миграции предшественников нейронов в области их окончательной дифференцировки (рис. 30), способствуя формированию правильной архитектоники мозга.
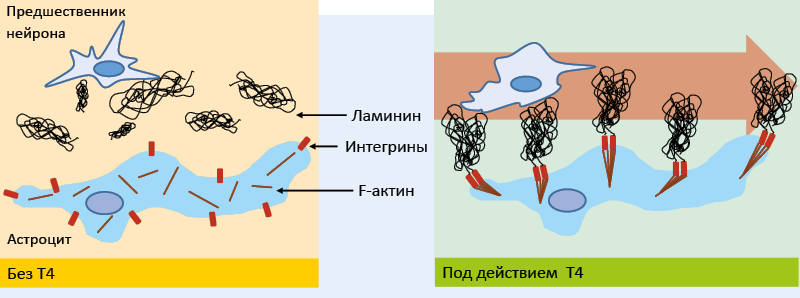
Рис. 30. Влияние Т4 на миграцию нейронов
Поступающий в мозг тироксин может оказывать в астроцитах негеномные эффекты, способствующие кластеризации молекул интегрина на мембране, что является необходимым условием для фиксации ламинина. Молекулы фиксированного на астроцитах ламинина создают выстилку для направленной миграции предшественников нейронов.
В процессе развития мозга тиреоидные гомоны влияют на:
- деление нейробластов;
- созревание и дифференцировку нейронов и олигодендроцитов;
- рост и ветвление дендритов;
- формирование цитоскелета астроцитов;
- выработку нейротрофинов и экспрессию их рецепторов;
- дифференцировку клеток гипофиза (лактотрофов и соматотрофов);
- процессы апоптоза;
- экспрессию генов, ответственных за миелинизацию.
От гормонов щитовидной железы зависит синтез G-белков, играющих важную роль в регуляции роста и дифференцировке нервных клеток (синхронизируют процессы дифференцировки, протекающие в мозге44). Дефицит гормонов щитовидной железы в период внутриутробного развития вызывает разнообразные нарушения развития нервной системы:
- задержку роста коры больших полушарий мозга и мозжечка;
- замедление роста тел нейронов переднего мозга;
- нарушение роста аксонов, дендритов и их миелинизации;
- наблюдается снижение ветвления дендритов клеток Пуркинье (рис. 31).
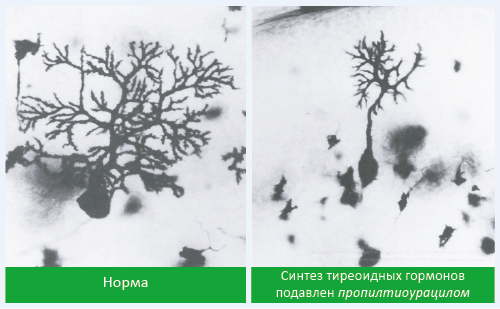
Рис. 31. Изменение структуры ганглионарных клеток под действием тиреоидных гормонов
Сходные закономерности наблюдаются в отсутствие функционирующих рецепторов тиреоидных гормонов, что продемонстрировано в животных моделях с нокаутом этих рецепторов:
- дефекты дифференцировки олигодендроцитов и астроцитов;
- дефекты нейрогенеза и ГАМК-эргических нейронов;
- нарушение синаптогенеза;
- дефекты мозжечка и клеток Пуркинье;
- тревожность;
- сниженная обучаемость;
- поведенческие дефекты спаривания;
- нарушение циркадных ритмов;
- глухота, дальтонизм.
Критические периоды, т.е. периоды наличия чувствительности клеток мозга к дифференцировочным эффектам тиреоидных гормонов, у человека захватывают период беременности45.
У взрослых особей тиреоидные гормоны также влияют на работу мозга:
- повышают уровень бодрствования и двигательную активность;
- поддерживают возбудимость ЦНС и ускоряют ассоциативное мышление;
- повышают активность ферментов, ускоряя включение аминокислот в белки;
- повышают интенсивность тканевого дыхания.
При повышении уровня тиреоидных гормонов выше нормы возбудимость ЦНС становится избыточной, а при снижении – недостаточной.
Регуляция метаболизма тиреоидными гормонами
Во взрослом организме тиреоидные гормоны, кроме поддержания возбудимости нервной системы, контролируют:
- работу сердца и почек;
- скорость поглощения кислорода;
- энергетический обмен;
- термогенез;
- метаболизм основных питательных веществ;
- продукцию ряда гормонов;
- репродуктивную функцию;
- моторику ЖКТ.
Гормоны щитовидной железы участвуют в поддержании нормальной функции дыхательного центра, увеличивают плотность β-адренорецепторов в сердце (стимулируют работу сердца, повышая чувствительность к катехоламинам46), скелетных мышцах, жировой ткани47, лимфоцитах. Помимо этого, Т3 и Т4 поддерживают работу ренин-ангиотензин-альдостероновой системы и прямо действуют на ряд транспортеров Na+ в почках, повышая почечный кровоток. Это, в свою очередь, ускоряет клубочковую фильтрацию и диурез.
Основными функциями тиреоидных гормонов во взрослом организме являются поддержание основного обмена и регуляция тканевого дыхания. В значительной мере их эффекты обеспечиваются изменениями в процессах окислительного фосфорилирования. Под действием тиреоидных гормонов:
- Растёт экспрессия термогенинов (увеличивают термогенез), встраивающихся во внутреннюю мембрану митохондрий и разобщающих окисление с фосфорилированием48;
- Увеличивается экспрессия и активация митохондриальных ферментов, отвечающих за окисление жиров и углеводов.
Действие гормонов щитовидной железы на окислительное фосфорилирование в митохондриях является главным механизмом стимулируемого ими калоригенеза.
Влияние тиреоидных гормонов на метаболизм основных нутриентов:
На метаболизм углеводов:
- Потенцируют действие инсулина на синтез гликогена и утилизацию глюкозы;
- Увеличивают скорость всасывания глюкозы в кишечнике;
- Увеличивают скорость захвата глюкозы жировой тканью и мышцами.
На метаболизм липидов:
- Усиливают липолиз;
- Стимулируют синтез и метаболизм холестерина49.
Поскольку влияние Т3 и Т4 на распад холестерина проявляется в большей степени, чем на синтез, при дефиците тиреоидных гормонов содержание ЛПНП50 и ЛПОНП51 в сыворотке возрастает. При избытке тироксина и трийодтиронина сывороточные концентрации этих веществ, напротив, снижаются.Тиреоидные гормоны являются объектом стимулирующего влияния гормона жировой ткани лептина52.
Информационные функции тиреоидных гормонов заключаются в стимуляции продукции СТГ и соматолиберина, Na+-уретических пептидов, гастрина, ГСПГ (снижая эффективность действия половых стероидов53).
Индекс атерогенности:
ИА = (общий холестерин – ЛПВП) / ЛПВП Интерпретация результата:
>3.5 – ↑ риск сердечно-сосудистых патологий
< 3.5 – норма
Так как ЛПОНП и ЛПНП являются проатерогенными (ЛПВП - напротив, антиатерогенны), повышение их концентраций у больных гипотиреозом ускоряет развитие атеросклероза, одной из причин гипертонической болезни.
И хотя тиреоидные гормоны способны снижать концентрации ЛПОНП и ЛПНП у здоровых людей, их применение в качестве гиполипидемических средств ограничено из-за выраженных побочных эффектов (раздражительность, бессонница, тремор, тахикардия и нарушения ритма сердца).
Рекомендуемая литература
1. «Элементарная фармакология», учебное пособие, В. В. Майский, 2005.
2. Agrawal N.K. (ed.) ThyroidHormones. – Second edition. – 2016, P. 385. ISBN-10: 953-51-0678-3
3. Arrojo E. Drigo R., FonsecaTL, Werneck-de-Castro JP, Bianco AC. // Role of the type 2 iodothyronine deiodinase (D2) in the control of thyroid hormone signaling. Biochim Biophys Acta. 2013 1830 (7): 3956-64.
4. Dentice M., Marsili A., Zavacki A., Larsen P.R., Salvatore D. // The deiodinases and the control of intracellular thyroid hormone signaling during cellular differentiation. – Biochim Biophys Acta. 2013 1830 (7): 3937-45.
5. Forhead A.J., Fowden A.L.// Thyroid hormones in fetal growth and prepartum maturation. – J Endocrinol. – 2014. – (3): R87-R103.
6. Hoermann R., Midgley J.E., Larisch R., Dietrich J.W. // Homeostatic Control of the Thyroid-Pituitary Axis: Perspectives for Diagnosis and Treatment. – 2015. – 6:177. ISBN-13: 978-953-51-0678-4
7. Moog NK, Entringer S, Heim C, Wadhwa PD, Kathmann N, Buss C. // Influence of maternal thyroid hormones during gestation on fetal brain development/-Neuroscience. 2015 pii: S0306-4522 (15) 00897-0.
8. Pappa T., Ferrara A.M., Refetoff S.// Inherited defects of thyroxine-binding proteins.– Best Pract Res Clin Endocrinol Metab. 2015 29 (5): 735–47.
9. Pascual A., Aranda A.// Thyroid hormone receptors, cell growth and differentiation.– Biochim Biophys Acta. 2013 (7): 3908-16.
10. Senese R., Cioffi F., de Lange P., Goglia F., Lanni A. Thyroid: biological actions of ‘nonclassical’ thyroid hormones. – J Endocrinol. 2014.– 221 (2):R1-12.
11. Sirakov M., Skah S., Nadjar J., Plateroti M.// Thyroid hormone’s action on progenitor/stem cell biology: new challenge for a classic hormone? – Biochim Biophys Acta. 2013 1830 (7):3917-27.
12. Skeaff SA. // Iodine deficiency in pregnancy: the effect on neurodevelopment in the child. – Nutrients. 2011.– 3 (2): 265-73.
38 Тиреотропин Рилизинг Гормон.
39 Уменьшает плотность рецепторов.
40 Представители семейства ФСГ: ЛГ, ФСГ, ХГч и ТТГ.
41 Активная аденилатциклаза (АЦ) синтезирует цАМФ.
42 Активная фосфолипаза С синтезирует IP3, увеличивающий [Са2+] в цитоплазме.
43 Глобулин, Связывающий Половые Гормоны.
44 Например, рост аксонов и их миелинизацию.
45 У грызунов критический период занимает вторую часть беременности и ранний постнатальный период.
46 Увеличивают сердечный выброс, частоту и силу сердечных сокращений.
47 β3-адренорецепторы обеспечивают липолитическое действие катехоламинов.
48 Термогенины (разобщающие белки) – ионные каналы, пропускающие Н+ по градиенту концентраций. При этом энергия градиента Н+ высвобождается в виде тепла.
49 В основном метаболизм.
50 ЛипоПротеины Низкой Плотности – переносчики холестерина к периферическим тканям.
51 ЛипоПротеины Очень Низкой Плотности – переносчики триглицеридов к периферическим тканям.
52 Гормон жировой ткани.
53 Связывая половые гормоны, уменьшает концентрацию их свободной формы в крови.