

Тканевые уровни, определяющие специфику и силу гормонального сигнала
Структуры, участвующие в формировании, передаче и реализации гормонального сигнала, можно разделить на:
- определяющие специфику сигнала:
– структура гормона;
– структура рецептора;
– активируемый каскад внутриклеточных посредников;
- регулирующие силу сигнала.
Специфика сигнала формируется, с одной стороны,структурой гормона (определяется продуцирующей клеткой/железой), с другой стороны – его рецепторным аппаратом (определяется тканью-мишенью), связывающим лиганды определенной структуры/класса/надсемейства. Однако в разных мишенях даже одинаковые рецепторы способны вызывать различные эффекты благодаря активации разных внутриклеточных каскадов.Эти механизмы позволяют получать тканеспецифичный ответ на каждое сигнальное соединение.
Спектр тканей-мишеней индивидуален для каждого гормона. И в зависимости от интенсивности воздействия гормона они подразделяются на: гормонзависимые, т.е. атрофирующиеся без воздействия гормона (напр., простата) и гормончувствительные, где гормон регулирует ряд метаболических/ростовых процессов, но не определяет жизнеспособность органа.
В регуляции силы гормонального сигнала участвуют гормонпродуцирующие элементы, системы транспорта и метаболизма сигнальных соединений, а также уровень рецепторов. Гормонпродуцирующая активность клеток обычно регулируется прямыми и обратными связями между гормонами одной вертикальной2 или горизонтальной3 оси. Концентрация гормонсвязывающих транспортных белков, экспрессия рецепторов и ферментов метаболизма находятся под более сложным мультигормональным контролем. Благодаря многоуровневой регуляции биодинамики, прямым/обратным связям, сила гормонального сигнала регулируется по замкнутому контуру, а концентрация вещества в крови поддерживается на постоянном уровне.
Эндокринный контур (рис. 9) в самом общем виде включает:
- Центральные звенья эндокринной оси, определяющие специфику и силу сигнала:
– эндокринная железа или гормонпродуцирующие клетки/ткани (продукция гормонов специфической структуры);
– чувствительные клетки/ткани (рецепторы, специфически узнающие конкретную группу гормонов).
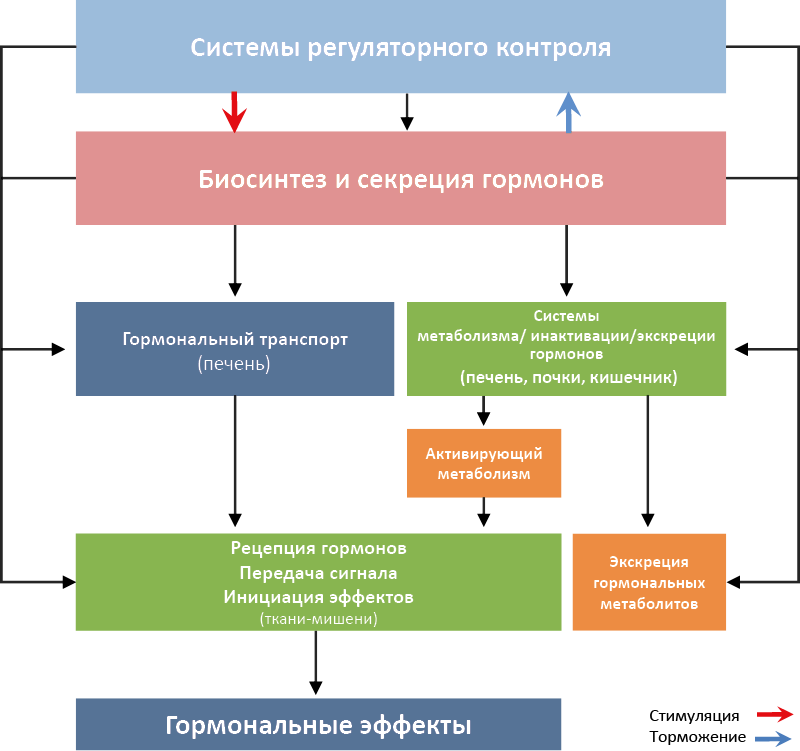
Рис. 9. Физиологическая организация эндокринного контура
- Органы, регулирующие силу гормонального сигнала:
– органы синтеза специфических транспортных белков, определяющих концентрацию свободного гормона в крови (преимущественно печень);
– ткани метаболизма/инактивации/экскреции гормонов (печень, почки и др.);
– регуляторные системы всех вышеперечисленных элементов контура (мозг, другие эндокринные железы, органы выделения метаболитов и т.д.).
Все компоненты этой системы – объекты гормональной регуляции. Их взаимодействие позволяет поддерживать концентрацию гормонов на относительно постоянном уровне, а системе работать по замкнутому контору (когда все элементы уравновешены). Благодаря этому система способна возвращаться в исходное положение после какого-либо возмущающего воздействия. Мутации препятствуют этому (Приложение 9).
Таким образом, концентрация гормона в крови – интегральный показатель активности эндокринного контура, объединяющая скорость продукции гормона и его метаболический клиренс (определяется скоростью метаболизма, наличием и концентрацией транспортных белков,плотностью рецепторов и имеющимися регуляторными воздействиями).
Элементы эндокринного контура
Гормоны
Гормоны – центральные звенья эндокринного контура, определяющие специфику и силу гормонального сигнала.У гормонов близкой структуры спектр биологической активности может пересекаться, поэтому наличие структурной гомологии является основанием для поиска сходства их эффектов.
В Приложении 1 представлены основные эндокринные железы и гормонпродуцирующие клетки разных уровней специализации, а также секретируемые ими соединения с гормональной и паракринной активностью. В Приложении 2 приведены примеры множественности тканевых источников гормонов, в Приложении 5 приведена классификация гормонов по структуре.
Транспортные гормонсвязывающие белки
Многие сигнальные соединения в кровотоке специфично и обратимо связываются своими транспортными белками, которые регулируют силу их гормонального сигнала. Гормон, связанный с транспортным белком, теряет биологическую активность и защищается от метаболизма. За счет этого снижается его метаболический клиренс и увеличивается период полужизни. Это позволяет создать депо гормона, буферирующее резкие изменения его концентрации в крови. В настоящее время открыто множество транспортных белков крови, преимущественно продуцируемых печенью (см. Приложение 6).
Название «транспортные» не полностью отражает функции этих белков (рис. 10). Кроме упомянутых функций депо и буфера гормональных колебаний, транспортные белки связаны с некоторыми физиологическими состояниями, например, беременностью. При беременности увеличивается продукция не только самих гормонов, но и их транспортных белков. Это позволяет защитить мать и плод от избыточных гормональных эффектов. Некоторые транспортные белки являются фрагментами рецепторов и регулируют доступность сигнального соединения для рецептора и, следовательно, эффективность проведения сигнала.

Рис. 10. Физиологическая роль транспортных белков
Рецепторы гормонов
Рецепторы определяют специфику и силу гормонального сигнала на уровне тканей-мишеней. Низкомолекулярные липофильные гормоны4 взаимодействуют с ядерными рецепторами (факторами транскрипции), которые связываются с ДНК в регуляторной области гормонзависимых генов и контролируют их экспрессию. Низкомолекулярные гидрофильные5 сигнальные соединения (катехоламины, простагландины), и белково-пептидные гормоны передают сигнал через мембранные рецепторы и последующие внутриклеточные каскады. Это приводит к фосфорилированию эффекторных белков или транскрипционных факторов (рис. 11).
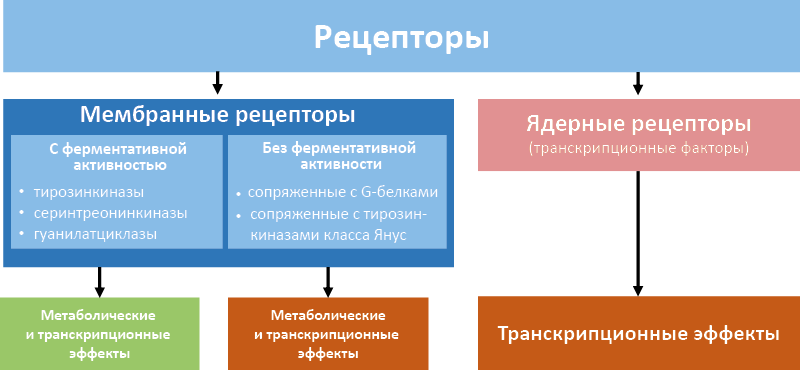
Рис. 11. Типы гормональных рецепторов
Мембранные рецепторы в общем виде делятся на:
- обладающие ферментативной активностью
– тирозинкиназы;
– серинтреонинкиназы;
– гуанилатциклазы;
- не обладающие ферментативной активностью;
– сопряженные с G-белками;
– сопряженные с тирозинкиназами класса Янус.
Эти рецепторы различаются типами внутриклеточных посредников и спектром последующих сигнальных каскадов. Каскады внутриклеточных посредников могут активироваться:
- представителями одного надсемейства рецепторов:
– напр. аденилатциклазный путь;
- представителями нескольких надсемейств рецепторов:
MAP-киназный каскад;
– обмен фосфатидилинозитолов и фосфоинозитидов.
В Приложении 7 представлены основные надсемейства рецепторов и взаимодействующие с ними гормоны.
Таким образом, для возникновения гормонального эффекта необходимо несколько последовательных этапов развития событий (рис. 12).
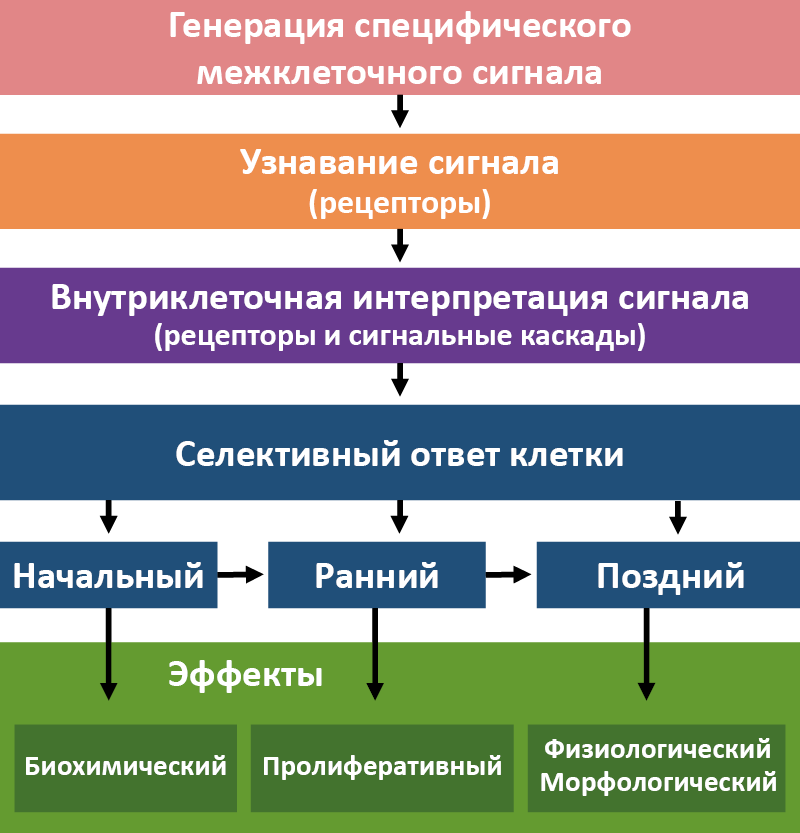
Рис. 12. Этапы инициации гормональных эффектов
Метаболические превращения гормонов
Сбалансированность эндокринного контура и содержание активного гормона в крови в большой мере определяются интенсивностью и направлениями метаболизма гормонов (рис. 13).
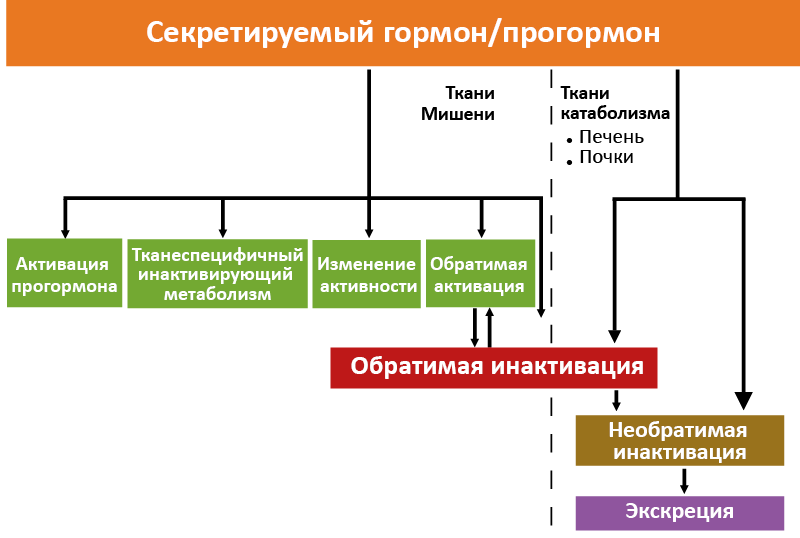
Рис. 13. Основные направления метаболизма гормонов
Гормоны могут не только инактивироваться в печени, почках и некоторых других органах (с последующими катаболизмом и экскрецией), но и претерпевать разнообразные метаболические превращения непосредственно в тканях-мишенях. Большое значение имеют варианты метаболизма, при которых ткань-мишень активно формирует тканеспецифичный ответ на гормон. Эти варианты можно подразделить на три группы:
- Изменение типа активности;
- Тканевая активация;
- Тканеспецифичная инактивация.
Метаболизм с изменением типа активности
Многие гормоны с разной биологической активностью структурно близки, и изменить их активность можно модификацией нескольких групп или фрагментацией белковой молекулы. Такие превращения может осуществлять ткань-мишень. Например, половая дифференцировка некоторых отделов мозга по мужскому типу происходит в результате превращения андрогенов в эстрогены ароматазой нейронов. Преобразование андрогенов в эстрогены происходит и в других тканях, например, жировой. Такое превращение вносит существенный вклад в продукцию эстрогенов в постменопаузу, когда их секреция яичниками снижается. У мужчин жировая ткань – равноправный участник поступления эстрогенов в системный кровоток.
Тканевая активация
Вторая группа связанных с тканью-мишенью преобразований–превращение в ней менее активного прогормона/гормона в более активное сигнальное соединение (напр., превращение тироксина в трийодтиронин в клетках-мишенях), что позволяет ткани-мишени активно регулировать силу гормонального сигнала.
Тканеспецифичная инактивация
Тканеспецифичный инактивирующий метаболизм – особый случай гормональных превращений, связанный со специфичностью узнавания гормонов рецептором. Хотя считается, что рецепторы почти абсолютно специфичны для своих гормонов, это не совсем так. Некоторые гормоны с близкой структурой, но разной биологической активностью могут с равным сродством перекрестно связываться с соответствующими рецепторами. Тканеспецифичный инактивирующий метаболизм позволяет ткани-мишени выбирать, какой гормон ей необходим для осуществления функции, а какой нужно инактивировать. Характерный пример – метаболическая система, существующая в почках. Рецепторы минералокортикоидов с одинаковым сродством связывают как минералокортикоид альдостерон, так и глюкокортикоид кортизол. В мозге кортизол действует и через глюкокортикоидные, и через минералокортикоидные рецепторы. В почках работает система инактивации кортизола – под действием 11β-гидроксистероиддегидрогеназы 2 типа кортизол инактивируется. Поэтому лишь альдостерон связывается с почечными минералокортикоидными рецепторами. Таким образом, в зависимости от ткани (сравните мозг и почки) действие кортизола через минералокортикоидные рецепторы может допускаться или не допускаться.
2 В случае осей гипоталамус-гипофиз-периферическая железа.
3 Напр., осей, связывающих гормоны-регуляторы углеводного и липидного обменов.
4 Стероидные, тиреодные, гормональные формы витамина D3 и производные витамина А, легко проникающие через билипидный слой.
5 Не способны проникнуть через клеточную мембрану.