

СТГ и регуляция его секреции
Центральное звено соматотропной оси – гормон роста86 аденогипофиза. Его гипоталамическими регуляторами являются соматолиберин (стимуляция продукции) и соматостатин (подавление продукции). Периферические регуляторы продукции СТГ – гормон желудка грелин (стимуляция) и ИФР-1 печени (подавление).Секреция СТГ контролируется несколькими системами обратной связи (рис. 59):
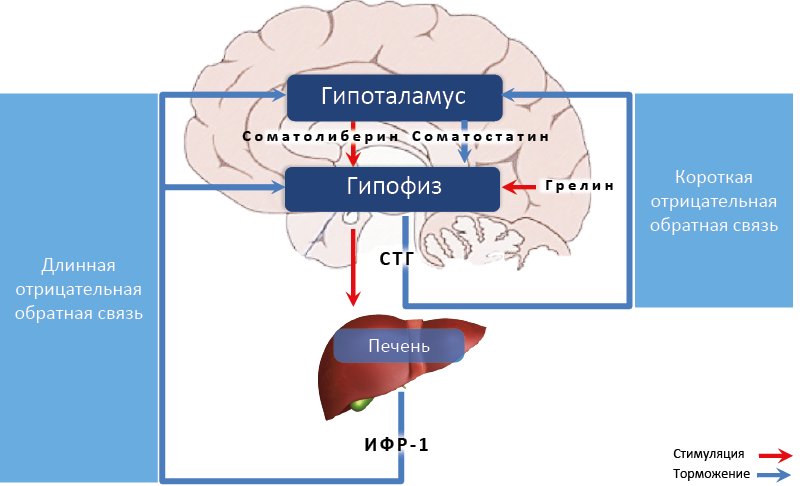
Рис. 59. Соматотропная ось
- Короткая отрицательная обратная связь
– подавление циркулирующим СТГ секреции соматолиберина (аркуатное ядро гипоталамуса) и стимуляция секреции соматостатина (перивентрикулярное ядро гипоталамуса).
- Длинная отрицательная обратная связь
– многоуровневое угнетение продукции СТГ циркулирующим ИФР-1 (растет в печени под действием СТГ) :
– в гипоталамусе уменьшает секрецию соматолиберина и стимулирует секрецию соматостатина;
– в аденогипофизе снижает секрецию СТГ, а также его чувствительность к грелину и соматолиберину, (ИФР-1 подавляет экспрессию их рецепторов в соматотрофах).
Соматостатин и соматолиберин регулируются ультракороткой обратной связью87. В регуляции секреции СТГ участвуют глюкокортикоиды и тиреодные гормоны (см. ниже). Выделение СТГ растет при голодании (на фоне возросшей продукции грелина) и при стрессе. Секреция СТГ носит пульсирующий характер, обусловленный эпизодической секрецией соматолиберина и соматостатина (контролируется супрахиазматическим ядром гипоталамуса), а эффективность его действия зависит от паттерна88 секреции. Эпизодическая секреция СТГ зависит от пола, возраста и сна:
- У женщин выше базальный уровень, частота секреции СТГ, но ниже её амплитуда.
- При старении секреция СТГ снижается.
- В фазу глубокого сна секреция СТГ всегда увеличивается.
Структура и механизмы действия гормонов соматотропной оси
СТГ – белковый гормон, структурно родственный пролактину и плацентарному лактогену89 (семейство пролактиноподобных белков). Наибольшей ростовой и анаболической активностями обладает СТГ, лактогенной – пролактин. У плацентарного лактогена лактогенные свойства и ростовая активность сопоставимы.
В крови значительная часть СТГ находится в комплексе с СТГ-связывающим белком, представляющим собой внеклеточный домен рецептора СТГ. Рецепторы пролактиноподобных белков (в т.ч. и СТГ) ассоциированы с тирозинкиназами класса Janus.
Соматолиберин – пептид, в состав его семейства входят такие сигнальные соединения как секретин, глюкагон, вазоактивный интестинальный пептид, гастроингибирующий пептид, орексины и др. Соматолиберин действует через рецепторы, сопряженные с G-белками, передавая сигнал преимущественно по аденилатциклазному пути (увеличивет [цАМФ]). Кроме стимуляции продукции СТГ, соматолиберин повышает потребление пищи и увеличивает продолжительность сна.
Соматостатин – пептид, структурно сходный с грелином и мотилином. Рецептор соматостатина также сопряжен с G-белками, но передает сигнал по фосфоинозитидному пути. Соматостатин является статином не только для СТГ, он снижает продукцию пролактина, инсулина и глюкагона, гормонов желудочно-кишечного тракта.
Грелин – пептидный гормон, синтезируемый в желудке и тонком кишечнике, а также, в небольшом количестве, в аркуатном ядре гипоталамуса. Он действует через свой рецептор, сопряженный с Gs-белком, индуцируя образование цАМФ и мобилизацию Ca2+. Кроме СТГ стимулирует продукцию пролактина, АКТГ, КРГ, вазопрессина, снижает продукцию соматостатина и негативно действует на гормоны гонадотропной оси.
Инсулиноподобный фактор роста 1 (ИФР-1) – одноцепочечный пептидный гормон, вхдящий в состав семейства инсулина. ИФР-1 передает сигнал через рецепторные тирозинкиназы.Как гормон продуцируется печенью, поступает в системную циркуляцию и регулирует работу соматотропной оси по механизму отрицательной обратной связи. Как паракринный фактор вырабатывается многими тканями и стимулирует в них процессы пролиферации. В кровотоке связан со своим переносчиком – ИФР-связывающим белком 3 (продуцируется печенью). Другие ИФР-связывающие белки (1, 2, 4–6) вырабатываются местно и регулируют активность ИФР-1 на паракринном уровне. Кроме стимуляции роста и пролиферации обладает инсулноподобной активностью, участвует в выборе доминантного фолликула.
Функции СТГ
Действуя на чувствительные ткани, СТГ (рис. 60):
- стимулирует рост организма:
– линейный;
– внутренних органов;
- оказывает анаболическое действие;
- оказывает нейротропное действие;
- вызывает липолиз;
- вызывает гипергликемию;
- стимулирует секрецию ИФР-1 печенью90.
- регулирует стрессорный ответ
СТГ влияет на некоторые типы клеток не только самостоятельно, но и с помощью ИФР-1, который дополняет и усиливает его действие (рис. 60).
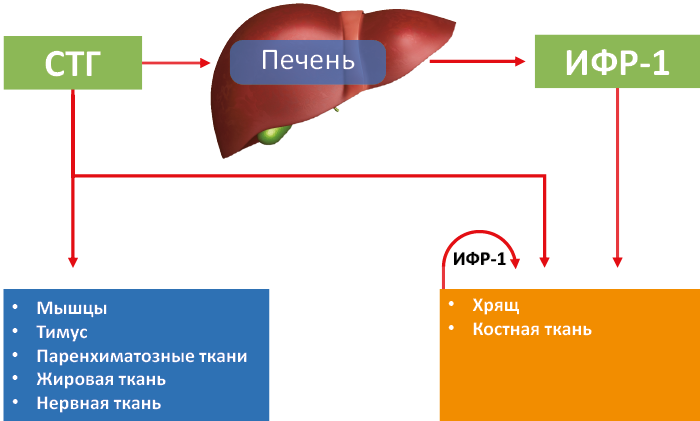
Рис. 60. Спектр СТГ-чувствительных тканей
Ростовые эффекты соматотропного гормона
Действуя на трубчатые кости, СТГ обеспечивает линейный рост организма. Этот эффект складывается из нескольких составляющих:
- СТГ прямо влияет на ранние стадии дифференцировки остеоцитов.
- Через продукцию ИФР-1 в костях, СТГ действует на поздние стадии дифференцировки этих клеток.
- Совместно с системным и местным ИФР-1 СТГ стимулирует пролиферацию остеобластов.
Анаболический эффект СТГ обеспечивается за счёт увеличения синтеза белка и активной утилизации жира в качестве источника энергии (а/к сохраняются для белкового синтеза). Активный белковый синтез достигается посредством:
- прямого геномного действия СТГ;
- увеличения захвата аминокислот мышцами и печенью.
СТГ стимулирует синтез белка в соединительной ткани, печени, мышцах, обуславливая их рост во время развития организма и анаболизм во взрослом периоде. СТГ, ИФР-1 и андрогены являются основными стимуляторами роста мышечной массы во взрослом организме.Выключение каждого из элементов соматотропной оси приводят к карликовости (рис. 61).

Рис. 61. Мутации соматотропной оси и вызываемые ими заболевания
Метаболические эффекты гормона роста
Под действием СТГ азотистый баланс91 сдвигается в положительную сторону. Так как аминокислоты активно поглощаются тканями и используются преимущественно в синтезе белка (анаболизме), концентрации аминокислот и мочевины92 в крови снижаются. Концентрация глюкозы, напротив, растёт из-за снижения чувствительности к инсулину и утилизации углеводов. Концентрация свободных жирных кислот в крови также увеличивается в результате усиленной утилизации жировых депо.
При стрессе СТГ выполняет сразу две функции: с одной стороны, быстро повышает глюкозу крови, с другой – уравновешивает катаболический эффект глюкокортикоидов.
Взаимосвязь СТГ и других гормонов
Кроме саморегуляции соматотропной оси СТГ способен изменять секрецию других сигнальных соединений (рис. 62). Например, увеличивать продукцию прогестерона, действуя прямо и через ИФР-1.
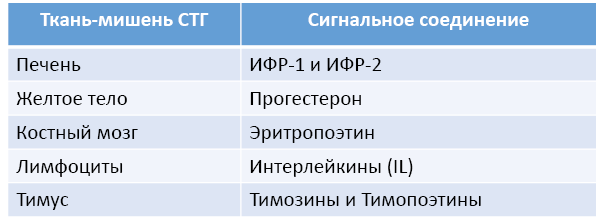
Рис. 62. Стимулирующее влияние СТГ на продукцию некоторых
сигнальных соединений эффекторными тканями
Другие гормоны в регуляции роста
Инсулин – помимо прочего является анаболическим гормоном и стимулирует рост. Ростовые эффекты особенно выражены в эмбриогенезе, однако клинически значимы и во взрослом организме (в отличие от СТГ он увеличивает жировые депо). В настоящее время применение в качестве анаболического средства ограничено из-за риска гипогликемии и ожирения.
Глюкокортикоиды – являются катаболическими гормонами (снижают синтез белка) для всех тканей кроме печени, поэтому, в целом, негативно влияют на рост. Однако их эффект зависит от текущей концентрации в крови, в низких дозах прямая стимуляция ими продукции СТГ превышает их катаболические эффекты.
Глюкокортикоиды обеспечивают:
- катаболизм всех тканей (кроме печени);
- продукцию печенью ИФР-1;
- дифференцировку соматотрофов гипофиза;
- прямую стимуляцию гена СТГ;
- подавление экспрессии гена грелина;
- снижение концентрации СТГ-РГ, но увеличение экспрессии его рецептора;
- увеличение продукции соматостатина, но снижение плотности его рецепторов.
Тиреоидные гормоны – оказывают стимулирующее влияние на рост, хотя их влияние на различные компоненты соматотропной оси разнонаправлено:
- снижение секреции СТГ-РГ, но увеличение экспрессии его рецептора;
- увеличение экспрессии гена грелина;
- стимуляция дифференцировки соматотрофов;
- стимуляция экспрессии гена СТГ.
Половые гормоны – регулируют рост, сначала вызывая его акселерацию, а затем – терминацию.
Рост каждой ткани определяет свой ансамбль гормонов, но особое положение занимают тимус и печень (рис. 63). В большинстве тканей анаболическими эффектами обладают СТГ, ИФР-1, инсулин и андрогены, а катаболическими – глюкокортикоиды. Однако в тимусе катаболическими эффектами обладают также и андрогены, вызывая его инволюцию при половом созревании. В печени все перечисленные гормоны оказывают анаболическое действие.
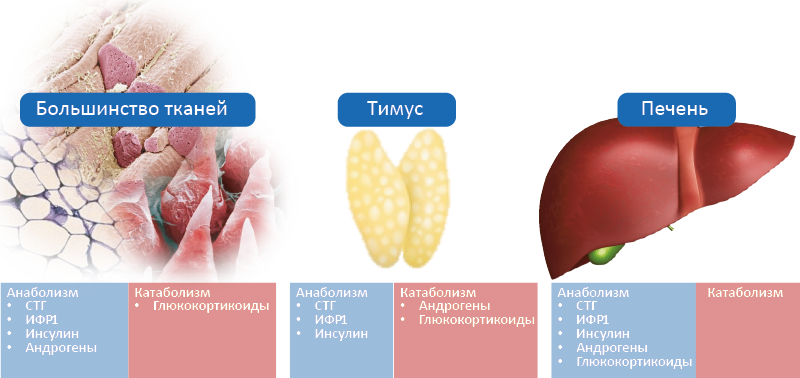
Рис. 63. Тканеспецифичные особенности мультигормональной регуляции роста
Вклад различных гормонов в регуляцию роста зависит от стадии онтогенеза (рис.64). В эмбриональном периоде рост обеспечивают преимущественно инсулин, ИФР-2 и ИФР-1. Главные регуляторы роста в постнатальном периоде – СТГ и ИФР-1.
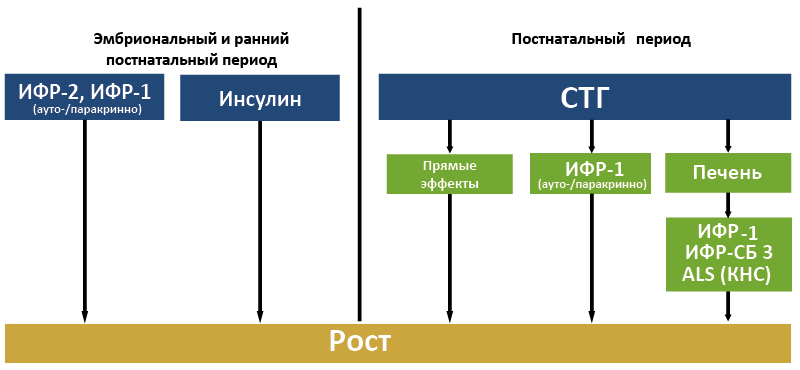
Рис. 64. Вклад различных гормонов в регуляцию роста на разных стадиях онтогенеза человека
Рекомендуемая литература
1. Baron J., Sävendahl L., De Luca F., Dauber A., Phillip M., Wit J.M., Nilsson O. // Short and tall stature: a new paradigm emerges.– Nat Rev Endocrinol. 2015 (12): 735-46.
2. Bowers M.D., Reynolds B.A. (auth.), Smith RG, Thorner MO (eds.) // Ghrelin in health and disease/-2012/-Humana Press. – P.290. – SBN: 9781617799020, 1617799025, 9781617799037, 1617799033
3. Clayton P.E., Gill M.S., Tillmann V., Westwood M. // Translational neuroendocrinology: control of human growth.– J Neuroendocrinol. 2014.– 26 (6):349-55. doi: 10.1111/jne.12156.
4. Darvin P., Joung Y.H., Yang Y.M.// JAK2-STAT5B pathway and osteoblast differentiation.– JAKSTAT. 2013.– 2 (4):e24931.
5. Hjortebjerg R., Frystyk J. // Determination of IGFs and their binding proteins.– Best Pract Res Clin Endocrinol Metab.– 2013.– 27 (6): 771-81.
6. Junnila R.K., List E.O., Berryman D.E., Murrey J.W., Kopchick J.J. // The GH/IGF-1 axis in ageing and longevity.– Nat Rev Endocrinol.– 2013.– 9 (6): 366-76.
7. Kaji H. // Interaction between Muscle and Bone.– J Bone Metab. 2014 21 (1):29-40.
8. Kasukawa Y., Miyakoshi N., Mohan S. // The anabolic effects of GH/IGF system on bone.– Curr Pharm Des. 2004.– 10 (21):2577-92.
9. Oberbauer A.M.// The influence of growth hormone on bone and adipose programming.– Adv Exp Med Biol. 2014.– 814: 169-76.
10. Theodoropoulou M., Stalla G.K. // Somatostatin receptors: from signaling to clinical practice.– Front Neuroendocrinol. 2013/-34 (3): 228-52.
11. Vottero A., Guzzetti C., Loche S.// New aspects of the physiology of the GH-IGF-1 axis.– Endocr Dev. 2013; 24:96-105.
12. Wang Y., Bikle D.D., Chang W. // Autocrine and Paracrine Actions of IGF-I Signaling in Skeletal Development.– Bone Res. 2013.– 1 (3): 249-59.
13. Yakar S., Isaksson O.// Regulation of skeletal growth and mineral acquisition by the GH/IGF-1 axis: Lessons from mouse models. – Growth Horm IGF Res. 2015/-pii: S1096-6374 (15) 30031-9.
86 Соматотропный гормон (СТГ).
87 Сами регулируют активность своих клеток-продуцентов.
88 Параметр, объединяющий концентрацию гормона и динамику её изменения.
89 Хорионический соматомаммотропин.
90 СТГ может также вызывать локальное образование ИФР-1 в различных органах и тканях.
91 Суточное поглощение азотсодержащих соединений превышает их экскрецию.
92 Продукт катаболизма аминокислот.