

Фосфатные соли113 Ca2+ составляют основной минеральный компонент костной ткани. Ca2+ необходим для сокращения мышечных волокон, внутриклеточного траффика и экзоцитоза везикул (включая синаптические пузырьки). Этот ион является внутриклеточным посредником многих сигнальных соединений, участвует в процессах деления и апоптоза. Концентрация Ca2+ в крови поддерживается на постоянном уровне с высокой точностью за счет действия ряда гормонов, наиболее значимыми из которых являются паратгормон, кальцитонин и гормональная форма витамина D (кальцитриол).
Паратгормон
Паратгормон (паратиреоидный гормон, ПТГ) – полипептид клеток паращитовидных желез, увеличивающий концентрацию Ca2+ и уменьшающий концентрацию фосфатов в крови.
Регуляция продукции
Между ПТГ, ионами Ca2+ и гормональной формой витамина D существуют реципрокные регуляторные связи (рис. 68): увеличение секреции паратгормона повышает содержание Ca2+ в крови и ускоряет синтез кальцитриола в почке; по механизму отрицательной обратной связи повышенные концентрации Ca2+ 114и кальцитриола угнетают продукцию паратгормона. Это происходит в результате угнетения экспрессии гена ПТГ и/или торможения его секреции клетками паращитовидных желез. Рост концентрации паратгормона приводит также к снижению концентрации фосфатов в крови. Секреция паратгормона снижается после приема пищи с высоким содержанием Ca2+.
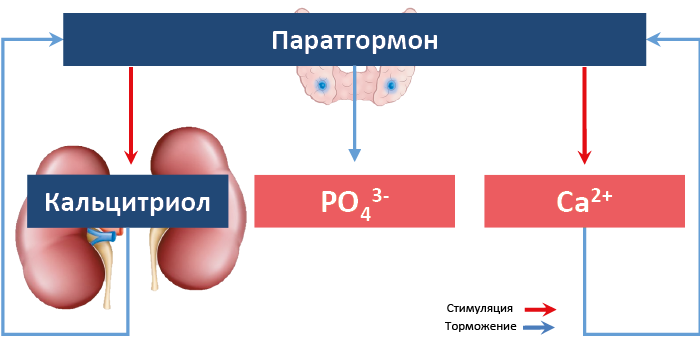
Рис. 68. Прямые и обратные связи в регуляции продукции паратгормона
Рецепторы
Рецепторы паратгормона – белки надсемейства рецепторов, сопряженных с G-белками. Передача сигнала осуществляется преимущественно по аденилатциклазному пути за счет увеличения продукции цАМФ. Классические органы-мишени – остеобласты и клетки проксимальных канальцев нефрона, где экспрессируются рецепторы паратиреоидного гормона.
Функции
Костная ткань
Гиперкальциемический эффект паратгормона достигается за счет:
- угнетения работы остеобластов
- активации остеоцитов, остеокластов
- увеличении пула остеокластов
Это способствует резорбции кости и поступлению Ca2+ и фосфатов из костной ткани в кровь.Паратгормон также индуцирует рецепторы кальцитриола в костной ткани.
Кишечник
Паратгормон опосредованно усиливает всасывание Ca2+ в кишечнике за счет стимуляции биосинтеза гормональной формы витамина D в почке.
Почки
Паратиреоидный гормон, действуя в восходящем колене петли Генле нефрона, стимулирует реабсорбцию Ca2+ и угнетает реабсорбцию фосфатов, увеличивая их выведение с мочой.В проксимальных канальцах нефрона он индуцирует экспрессию 1α-гидроксилазы, ключевого фермента биосинтеза кальцитриола, и уменьшает количество мРНК 24-гидроксилазы, катализирующей образование малоактивного метаболита витамина D. Суммарный эффект паратгормона на кости и почки сводится к увеличению концентрации Ca2+ и снижению концентрации фосфата во внеклеточной жидкости. Разница их концентраций обеспечивает дополнительную диссоциацию фосфатных солей Ca2+ в костях. В почках паратгормон также ингибирует реабсорбцию бикарбонатов, натрия и жидкости.
Частой причиной развития остеомаляции у женщин в постменопазе является дефицит эстрогенов. Эти и другие проявления менопаузы хорошо поддаются коррекции с помощью гормональной заместительной терапии. Для этого используют препараты с вариабельным содержанием эстрогенов и прогестинов, имитирующие естественные колебания этих гормонов во время менструального цикла.
Гормональная форма витамина D (Кальцитриол)
Гормональная форма витамина D (1α,25-дигидроксивитамин D, кальцитриол) синтезируется последовательным гидроксилированием холекальциферола (витамина D): сначала в печени по 25-му положению с образованием малоактивного 25-гидроксивитамина D, затем в почке по 1α-положению с образованием 1α,25-дигидроксивитамина D.
Регуляция продукции
Регуляция образования кальцитриола осуществляется преимущественно на уровне 1α-гидроксилазы проксимальных почечных канальцев. Биосинтез этого фермента стимулируется паратгормоном и/или пониженным содержанием фосфата крови. Активный гормон не только ингибирует свою продукцию по механизму отрицательной обратной связи (подавляя продукцию ПТГ и экспрессию 1α-гидроксилазы), но и усиливает собственную деградацию, стимулируя транскрипцию 24-гидроксилазы.
Витамин D и его гидроксилированные метаболиты, включая кальцитриол, транспортируются в комплексе с транспортным белком крови, связывающим витамин D (транскальциферином). Причем образовавшийся в печени 25-гидроксивитамин D обладает наивысшим сродством к этому белку. В отличие от большинства гормонсвязывающих белков, витамин D-связывающий белок не регулирует биодоступность, а направленно транспортирует 25-гидроксивитамин D из первичной мочи в проксимальные трубочки для 1α-гидроксилирования и активации.
Рецепторы
Рецептор кальцитриола относится к надсемейству ядерных рецепторов и экспрессируется в классических органах-мишенях кальцитриола (кишечник, кости, почки, паращитовидные железы) и многих других.
Функции
Кальцитриол способствует поддержанию постоянной концентрации внеклеточного Ca2+, действуя на кишечник, кости, почки и паращитовидные железы.
Кишечник
Кальцитриол – основной стимулятор всасывания Ca2+ и фосфатов в тонком кишечнике. Регуляция всасывания под действием кальцитриола может затрагивать различные механизмы (облегчённую диффузию, везикулярный транспорт) в зависимости от отдела кишечника.
Костная ткань
Кальцитриол стимулирует накопление Ca2+ в остеоцитах и остеобластах, участвует в развитии и минерализации костной ткани, ремоделировании костей (совместно с паратгормоном, потенциируя действие последнего).
Почки
Кальцитриол стимулирует реабсорбцию Ca2+ в дистальных почечных канальцах, снижая его экскрецию с мочой. Это происходит за счет увеличения экспрессии генов системы трансцеллюлярного транспорта Ca2+. Снижение выведения ионов Ca2+ почками приводит к повышению их концентрации в крови.
Паращитовидная железа
Кальцитриол подавляет транскрипцию гена паратгормона. Действуя на другие ткани, кальцитриол стимулирует секрецию кальцитонина С-клетками щитовидной железы и адреналина мозговым слоем надпочечников.
Кальцитонин
Кальцитонин – гипокальциемический и гипофосфатемический гормон, синтезируемый С-клетками щитовидной железы.
Регуляция продукции
Продукцию кальцитонина стимулируют гастрин, Ca2+, глюкагон и GLP115, секреция которых растет в ответ на прием пищи. Наибольшим эффектом обладает гастрин. Предполагают, что увеличение секреции кальцитонина в ответ на эти гормоны является защитой от пищевой гиперкальциемии. В этом проявляется его основная функция, поскольку удаление щитовидных желёз практически не меняет содержание Ca2+ и фосфата в крови. Повышение концентраций Ca2+ и кальцитриола в крови также стимулирует секрецию кальцитонина.
Рецепторы
Рецепторы кальцитонина – белки надсемейства рецепторов, сопряженных с Gs-белками. Cигнал передаётся по аденилатциклазному пути за счет увеличения продукции цАМФ.
Функции
В норме вклад кальцитонина в регуляцию уровня Ca2+ в крови относительно невелик и заключается в защите от пищевой гиперкальциемии, однако при аномалиях работы паращитовидной железы эффекты кальцитонина отчётливо видны. Это позволяет считать кальцитонин антагонистом паратгормона.
Костная ткань
Кальцитоин тормозит дифференцировку предшественников-остеокластов, ослабляет резорбцию костей и высвобождение из них Ca2+ и фосфатов.
Кишечник
Кальцитонин снижает трансцеллюлярный транспорт Ca2+ в кишечнике.
Почки
В почках кальцитонин подавляет реабсорбцию и Ca2+, и фосфата. Однако этот эффект выражен только при сверхфизиологических концентрациях гормона. Как и паратгормон, в почках кальцитонин усиливает экспрессию 1α-гидроксилазы, ключевого фермента синтеза кальцитриола.
Кроме вышеописанных гормонов, в регуляции гомеостаза Ca2+ участвуют глюкокортикоиды, тормозящие ремоделинг костей (подавляют дифференцировку остеобластов, всасывание Ca2+ кишечником и его реабсорбцию почками), эстрогены, увеличивающие прочность костей (стимулируют продукцию кальцитонина, работу остеобластов) и некоторые другие116. Взаимосязи гормонов, регулирующих отмен кальция представлены на рис. 69.
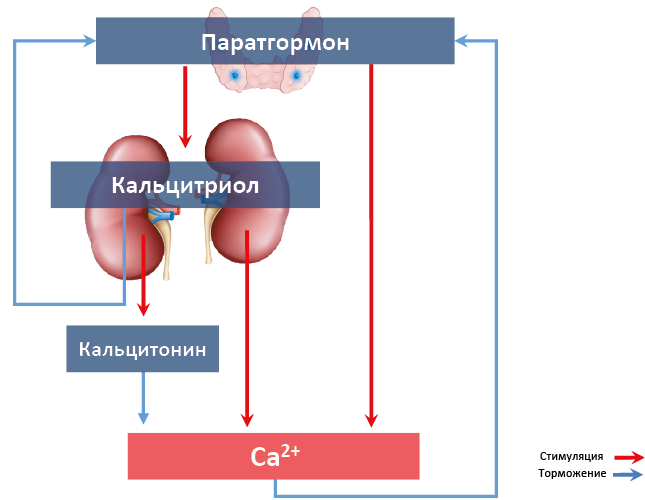
Рис. 69. Гормоны, влияющие на концентрацию Ca2+, и связи между ними
Рекомендуемая литература
1. Briot K., Roux C.//Glucocorticoid-induced osteoporosis. -RMD Open. -2015-V. 1 (1): e000014.
2. Cheloha R.W., Gellman S.H., Vilardaga J.P., Gardella T.J. // PTH receptor-1 signalling-mechanistic insights and therapeutic prospects. -Nat Rev Endocrinol. 2015-V.11 (12):712-24.
3. Hendy G.N., Canaff L. Calcium-sensing receptor, proinflammatory cytokines and calcium homeostasis Semin Cell Dev Biol. 2015- S1084-9521 (15) 30006-9.
4. Jeon U.S. // Kidney and calcium homeostasis. -Electrolyte Blood Press. -2008 -V.6 (2): 68-76.
5. Nagano N //Pharmacological and clinical properties of calcimimetics: calcium receptor activators that afford an innovative approach to controlling hyperparathyroidism. – Pharmacol Ther. 2006 -V.109 (3): 339–65.
6. Naot D., Cornish J. // The role of peptides and receptors of the calcitonin family in the regulation of bone metabolism. – Bone. 2008-V.43 ( 5): 813-8.
7. Taylor C.W., Tovey S.C. // From parathyroid hormone to cytosolic Ca2+ signals. – Biochem Soc Trans. 2012 -V.40 (1): 147-52.